
Основные органеллы цитоплазмы
У зеленой водоросли празиолы (Prasiola stipitata) обнаружена соматическая редукция — мейоз совершается в вегетативных клетках верхней части диплоидного гаметофита, при этом появляются участки гаплоидных клеток, в которых вслед за тем образуются гаплоидные гаметы (рис. 25, 4).
В цикле развития водорослей, лишенных полового размножения (сине-зеленые, криптофитовые и эвгленовые) или имеющих его в редких случаях (золотистые, желто-зеленые и динофитовые), наблюдаются только изменения в строении тела.
Поэтому применительно к таким водорослям принято говорить о цикломорфозе. Он может охватывать несколько поколений или ограничиваться периодом роста и развития одной особи. В наиболее резкой форме цикломорфоз выражен у гиеллы дернистой (Hyella caespitosa) из сине-зеленых водорослей и у гленодиниума Борге (Glenodinium borgei) из динофитовых.
Как циклы развития, так и цикломорфозы у водорослей отличаются большой пластичностью.
Их прохождение во многом определяется экологическими условиями. Поэтому они далеко пе всегда сопровождаются строго последовательным проявлением всех стадий. В зависимости от условий произрастания отдельные стадии и формы развития могут выпадать полностью (например, спорофит или гаметоспорофит и гаметофит) или, наоборот, существовать на протяжении нескольких поколений, с тем чтобы на период жизни одного поколения уступить место другой форме развития.
Строго упорядоченные циклы развития существуют у водорослей, стоящих на верхних ступенях эволюции
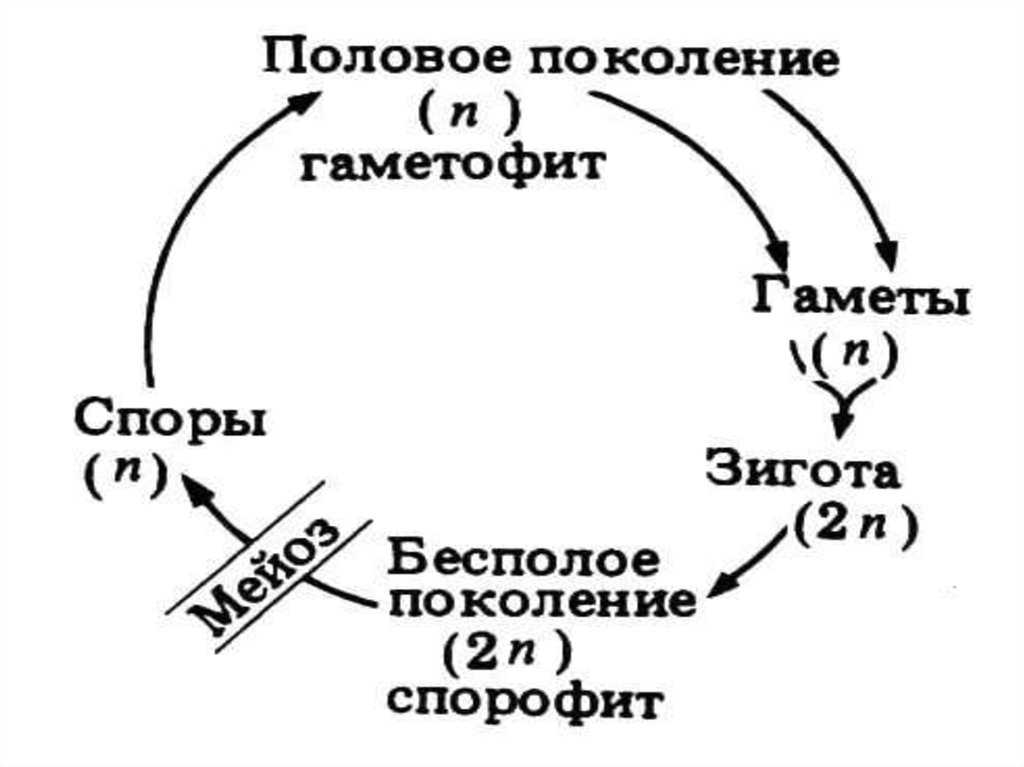
Соотношение диплоидной и гаплоидной фаз в жизненном цикле разных водорослей неодинаково.
В одних случаях прорастание зиготы сопровождается редукционным делением конуляционного ядра (зиготическая редукция) и развивающиеся при этом растения оказываются гаплоидными. Так, у многих зеленых водорослей (вольвокальные, большинство хлорококкальных, ко-нъюгатофициевые, харофициевые) зигота —единственная диплоидная стадия в цикле развития, вся вегетативная жизнь проходит у них в гаплоидном состоянии, они являются гаплонтами.
У других водорослей, наоборот, вся вегетативная жизнь осуществляется в диплоидном состоянии, а гаплоидная фаза представлена лишь гаметами, перед образованием которых и происходит редукционное деление ядра (гаметическая редукция). Зигота без редукционного деления ядра прорастает в диплоидный таллом. Эти водоросли— диплонты. Таковы многие зеленые водоросли, имеющие сифоновое строение, все диатомовые, из бурых — представители порядка фукальные.
У третьих редукционное деление ядра предшествует образованию зооспор или анланоспор, развивающихся, как правило, на диплоидных талломах (спорическая редукция). Эти клетки бесполого размножения вырастают в гаплоидные растения, размножающиеся только половым путем.
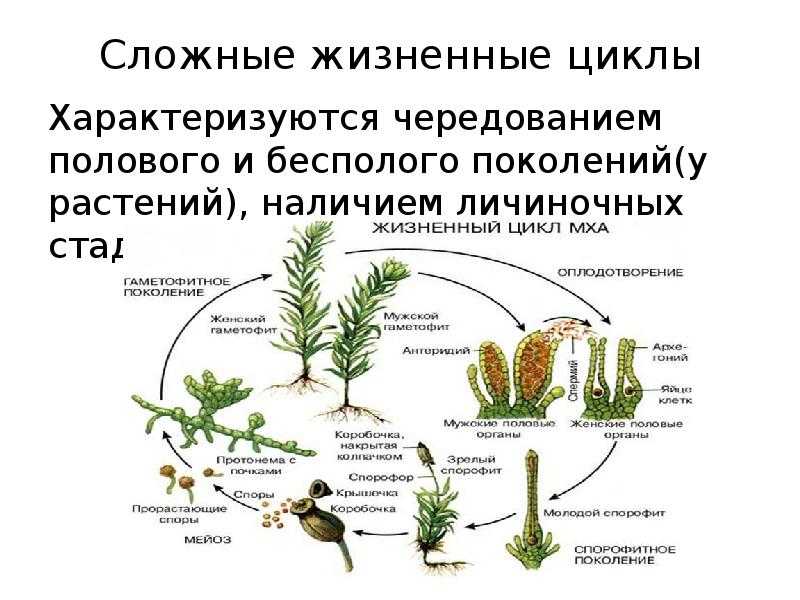
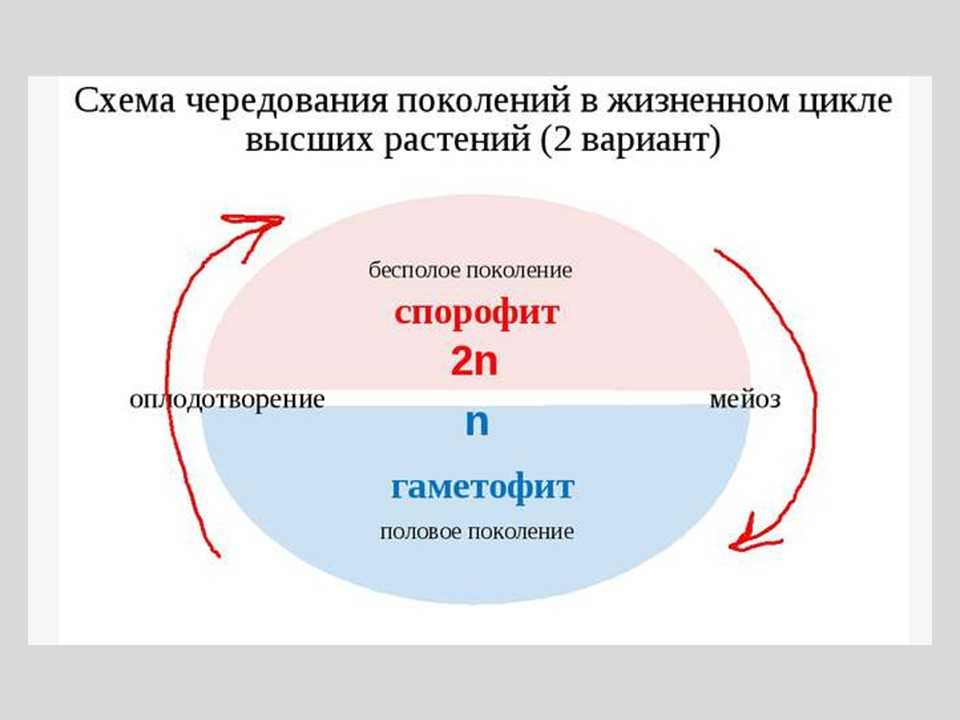
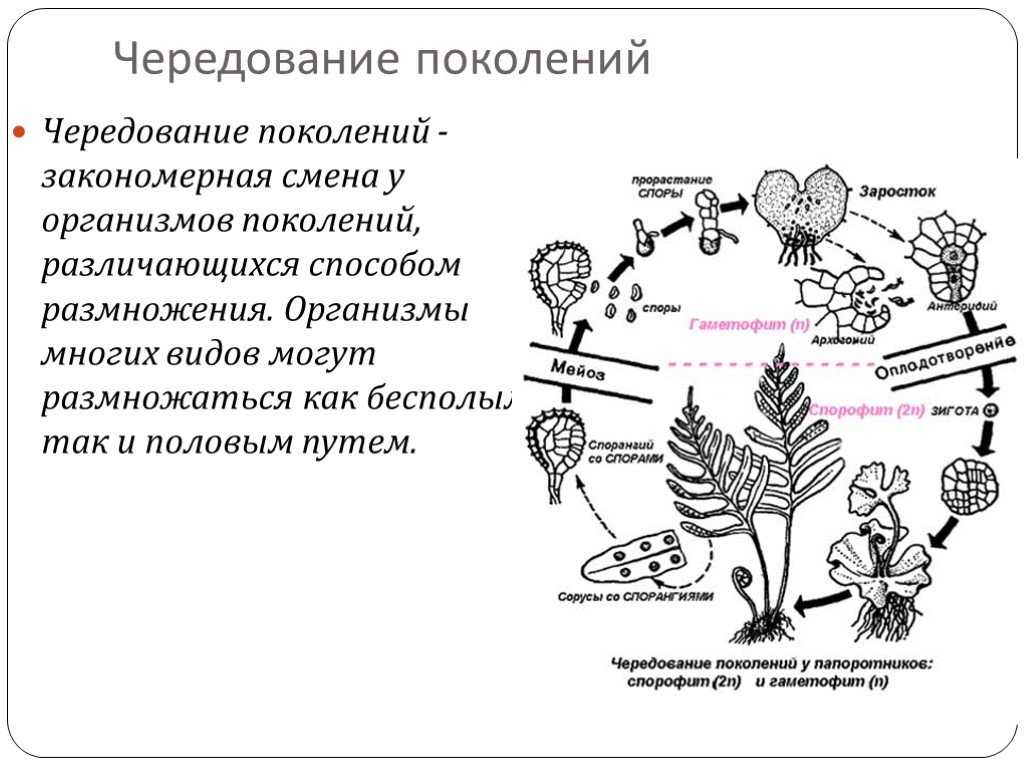
После слияния гамет зигота развивается в диплоидное растение, несущее только органы бесполого размножения. Таким образом, у этих водорослей имеет место чередование форм развития (генераций): диплоидного бесполого спорофита и гаплоидного полового гаметофита.
Оба поколения могут быть одинаковы морфологически (изоморфная смена генераций) или же резко различны по внешнему виду (гетероморфная смена генераций).
Изоморфная смена генераций характерна для морских видов ульвы, энтероморфы, кладофоры, хетоморфы из зеленых водорослей, для ряда порядков бурых и большинства красных водорослей (несколько усложненная), Гетероморфная смена генераций особенно распространена среди бурых водорослей, но встречается у зеленых и красных.
Половое и бесполое размножение: в чем разница
Существует два метода размножения: половой и бесполый. Половое размножение включает в себя такие фундаментальные процессы, как гаметогенез и оплодотворение.
Гаметогенез — это биологический процесс, представляющий собой деление клеток, в частности митоз и мейоз, для производства гамет (половых клеток). Он начинается с того, что первичные клетки делятся митотически, а затем каждая из них подвергается мейозу.
Мейоз — это форма деления клетки, при которой клетка проходит через два последовательных клеточных деления. В результате образуются четыре гаплоидные клетки. Затем каждая гаплоидная клетка проходит процесс созревания и становится полностью дифференцированной гаметой (половой клеткой).
У женщин зрелая гамета — это яйцеклетка. У мужчин зрелой гаметой является сперматозоид. Мужская гамета должна найти путь к яйцеклетке, чтобы соединиться с ней. У высших форм животных самец и самка, обычно одного вида, вступают в половой акт, чтобы сблизить гаметы для соединения.
Сперматозоид, найдя яйцеклетку, проникает в нее, чтобы их генетический материал объединился в один полный набор (диплоидный). Этот союз знаменует собой оплодотворение, и оплодотворенная яйцеклетка проходит серию митотических делений, чтобы дать начало эмбриону, состоящему из диплоидных клеток.
Оплодотворение яйцеклетки схематично:
Асексуальное размножение отличается от полового тем, что потомство было получено бесполым путем. Процесс отличается тем, что не включает в себя мейоз и оплодотворение. Более того, родительский организм производит потомство даже при отсутствии пары. Потомство, скорее всего, будет клоном родительского организма.
Этапы менструального цикла
Чтобы интерпретировать явления, сопровождающие физиологические репродуктивные процессы, и эффективно выявлять нарушения в репродуктивной системе, нужно понимать, как функционируют гипоталамус, гипофиз, яичники и матка в менструальном цикле.
Яичники — репродуктивные органы, выполняющие зародышевую и гормональную функцию. Патологическое состояние яичников, нарушающее любую из функций, приводит к бесплодию.
В цикле в яичнике происходит:
- образование яйцеклеток (зародышевая функция);
- секреция гормонов — (гормональная функция).
Развитие яйцеклеток в свою очередь связано с активностью гипофиза, секретирующего гормоны гонадотропины. Фолликулы развиваются в две фазы:
- независимо от гонадотропинов – под контролем местных факторов (до антральных пузырьков);
- под контролем гонадотропинов.
Гонадотропины отвечают за:
- развитие фолликула в гонадотропин-зависимой фазе;
- секрецию эстрадиола;
- завершение созревания яйцеклетки;
- овуляцию;
- образование желтого тела;
- поддержание выработки прогестерона и эстрадиола желтым телом.
В гонадотропинозависимой фазе цикла участвуют разные гормоны, производимые в гипоталамусе, гипофизе, яичниках (гипоталамо-гипофизарно-гонадная ось):
- гипоталамический гонадолиберин, регулирующий гонадотропин (ГнРГ) рилизинг-гормон;
- гонадотропины: лютеинизирующий гормон (ЛГ) и фолликулостимулирующий гормон (ФСГ);
- эстрадиол, отвечающий за разрастание слизистой оболочки матки;
- прогестерон, подготавливающий слизистую оболочку к имплантации плодного яйца.
Нарушения в гипоталамо-гипофизарно-гонадной оси приводят к нарушениям менструального цикла, плохому общему самочувствию и бесплодию.
Сосудистые и несосудистые растения
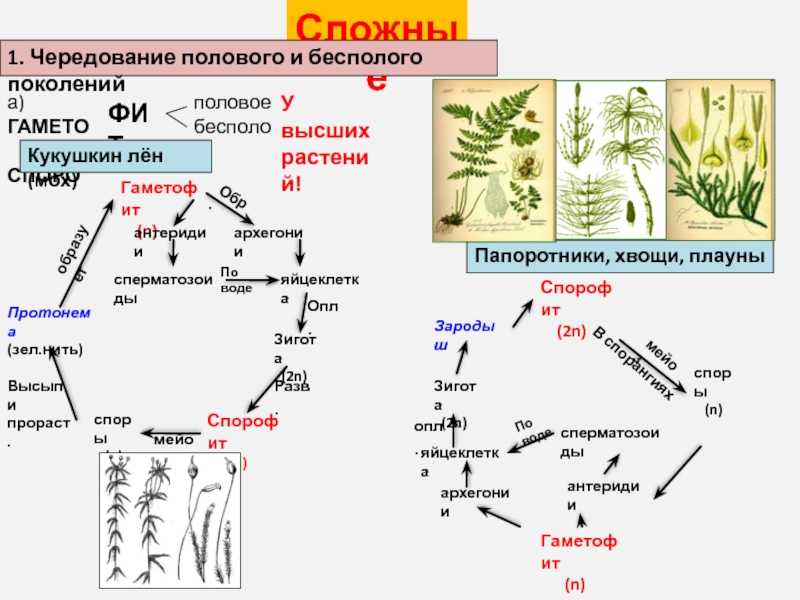
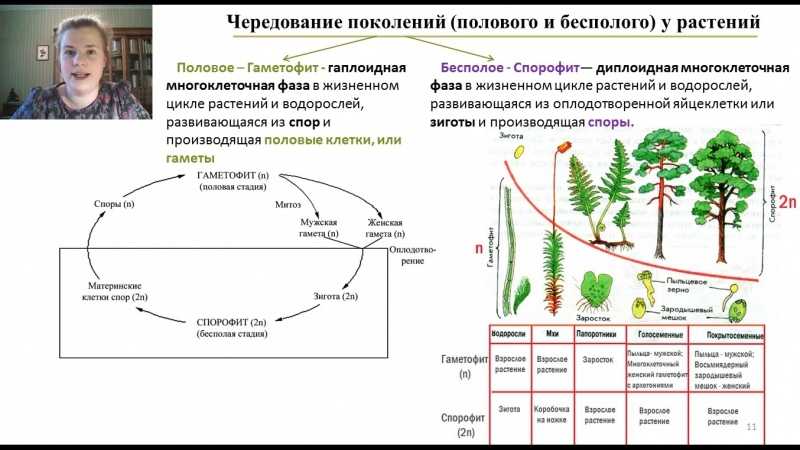
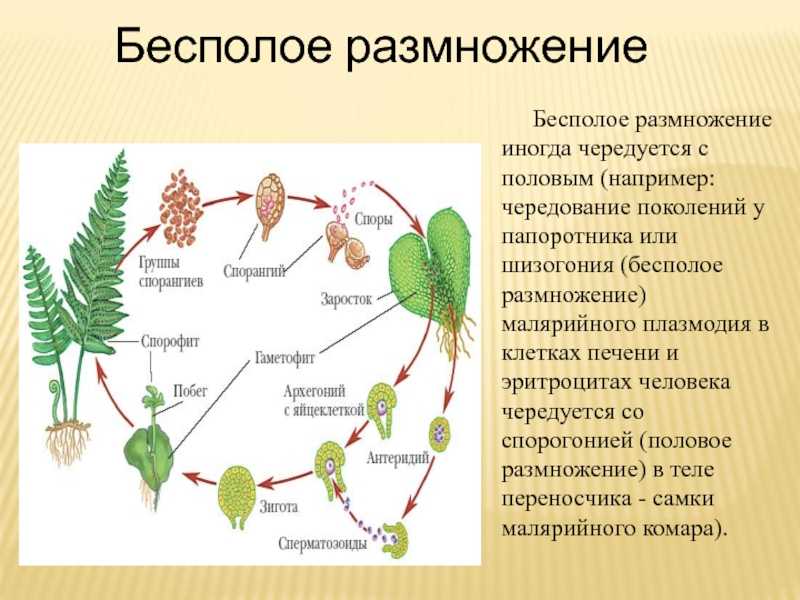
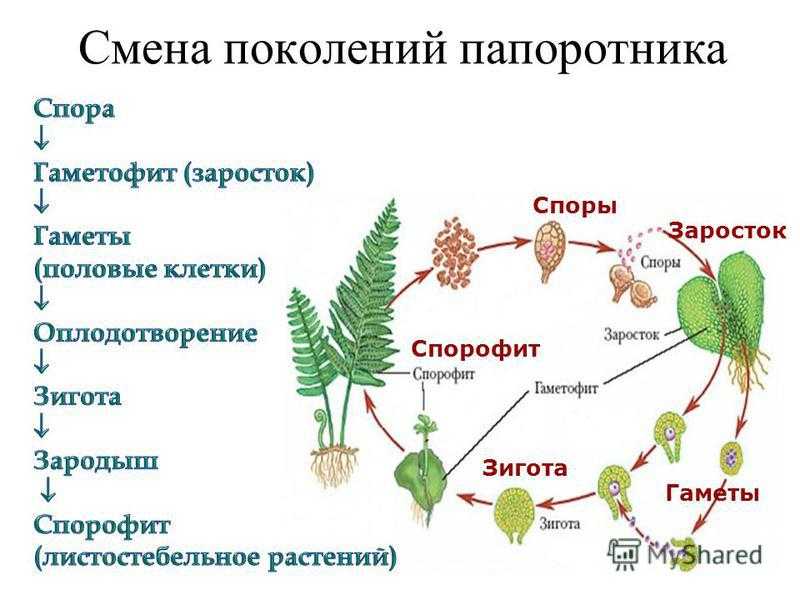
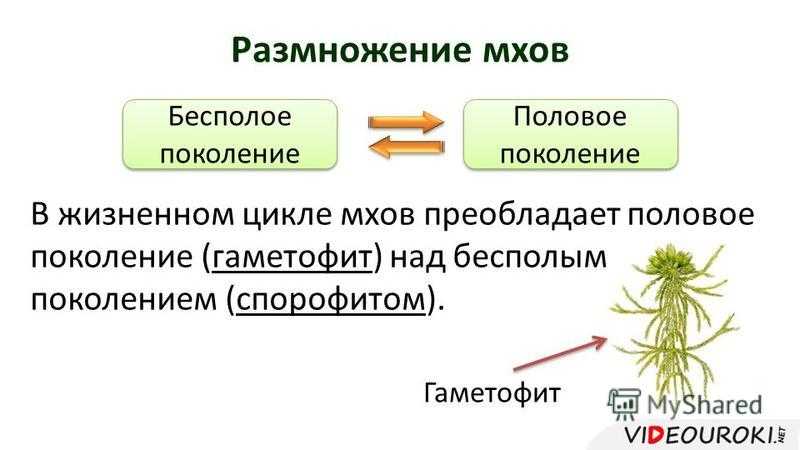
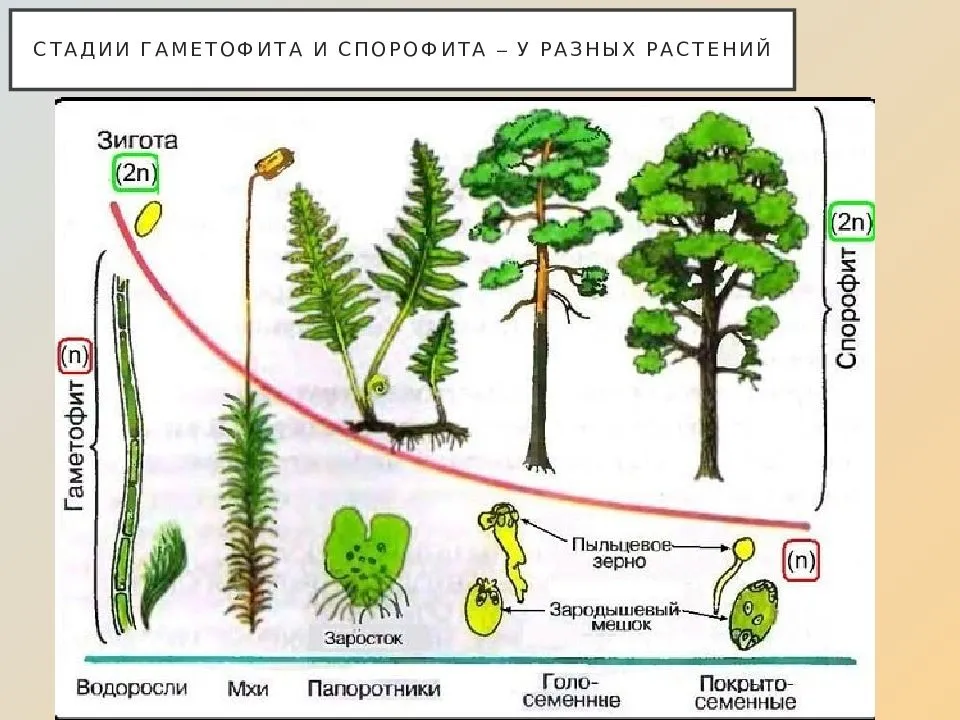
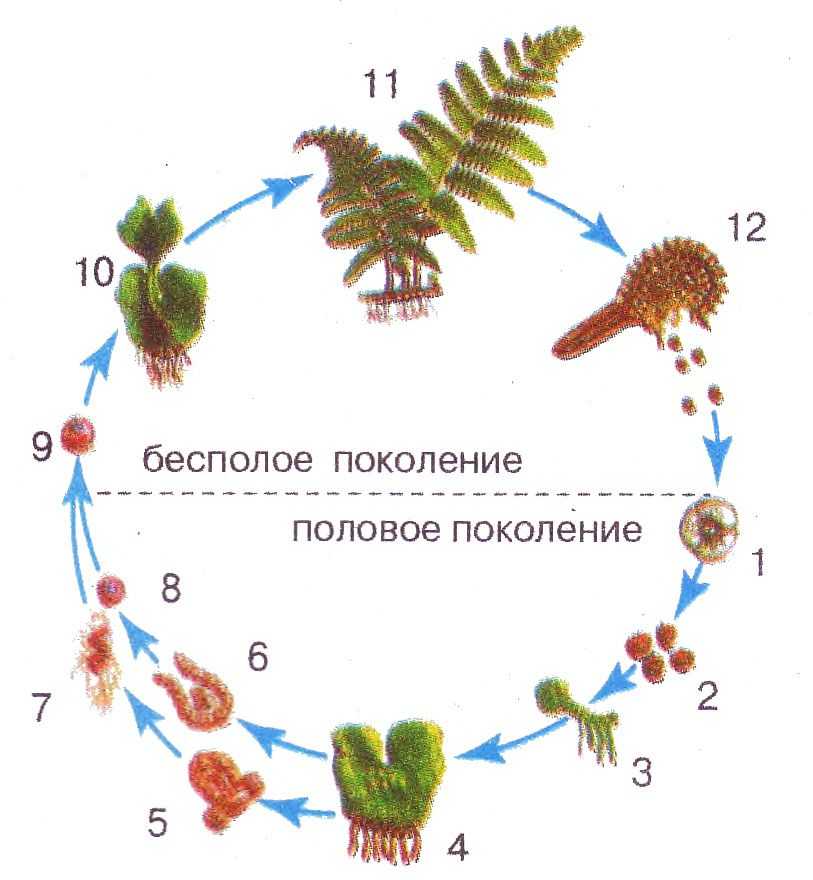
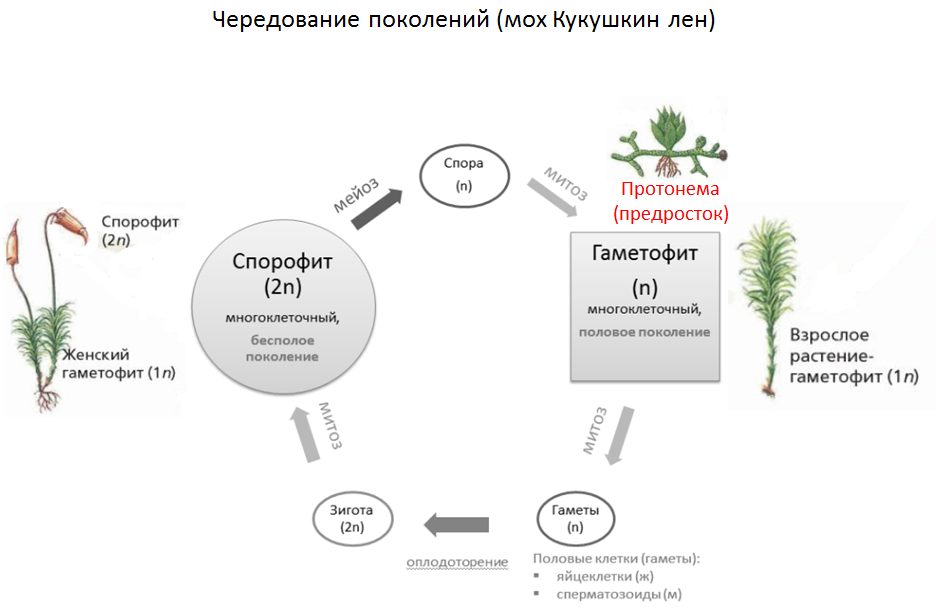
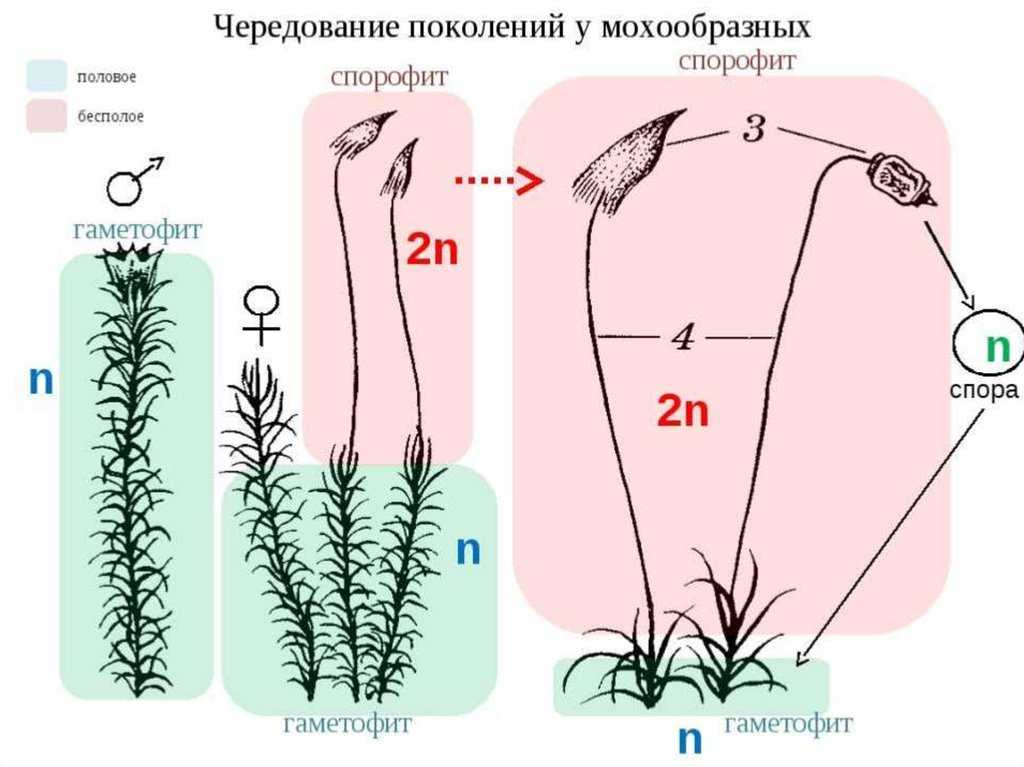
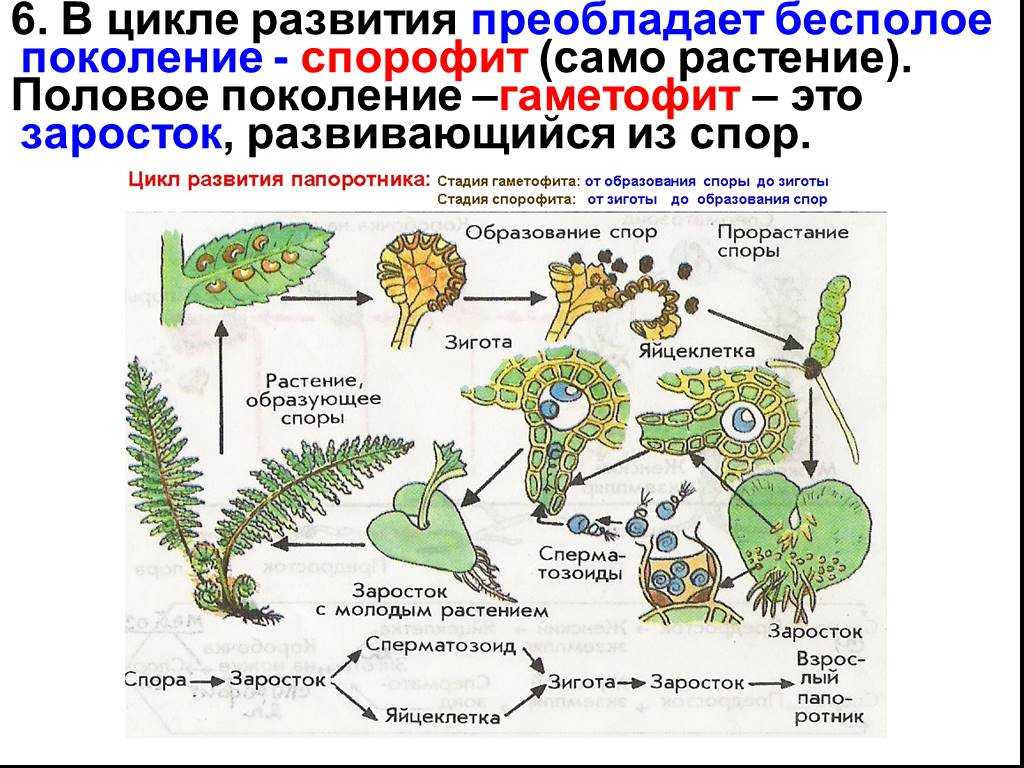
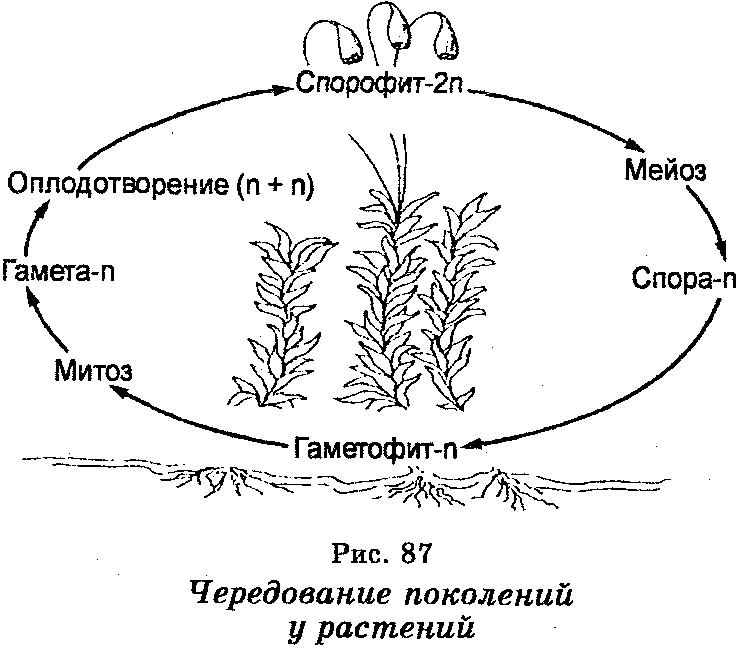
Чередование поколений наблюдается как у сосудистых, так и несосудистых растений. Сосудистые растения содержат систему сосудистой ткани, которая транспортирует воду и питательные вещества по всему телу растения. Несосудистые растения не имеют такой системы и нуждаются во влажных местах обитания для выживания. К ним относятся мхи, антоцеротовидные и печёночные мхи. Эти растения выглядят как зеленые маты растительности с выступающими из них стебельками. Первичной фазой жизненного цикла несосудистых растений является генерация гаметофитов. Фаза гаметофит состоит из зеленой мшистой растительности, а фаза спорофит состоит из удлиненных стеблей со спорангиями на концах.
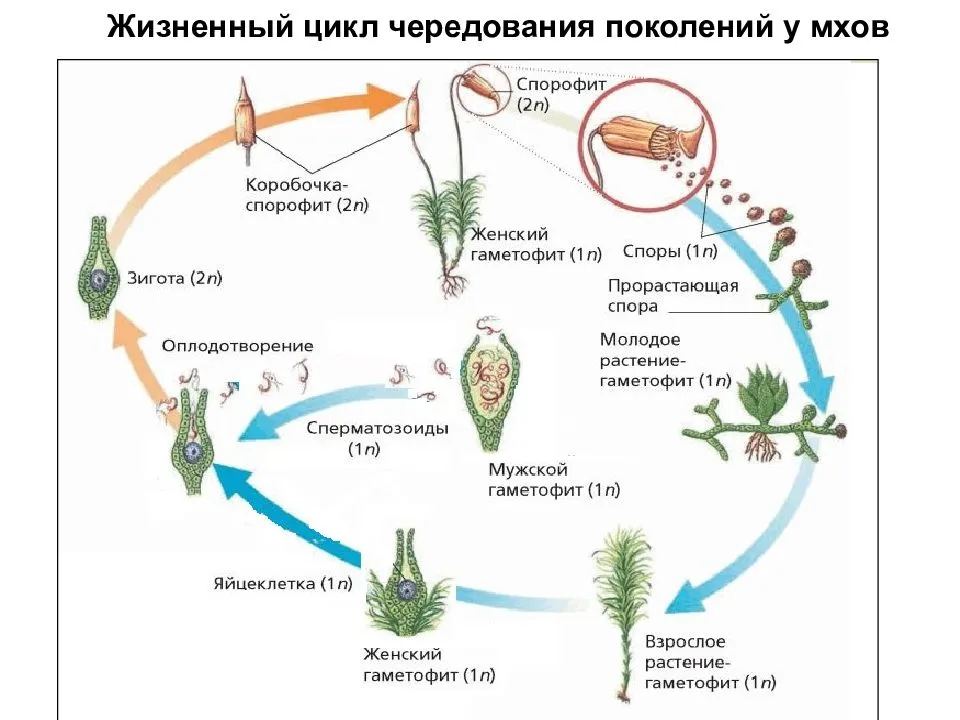
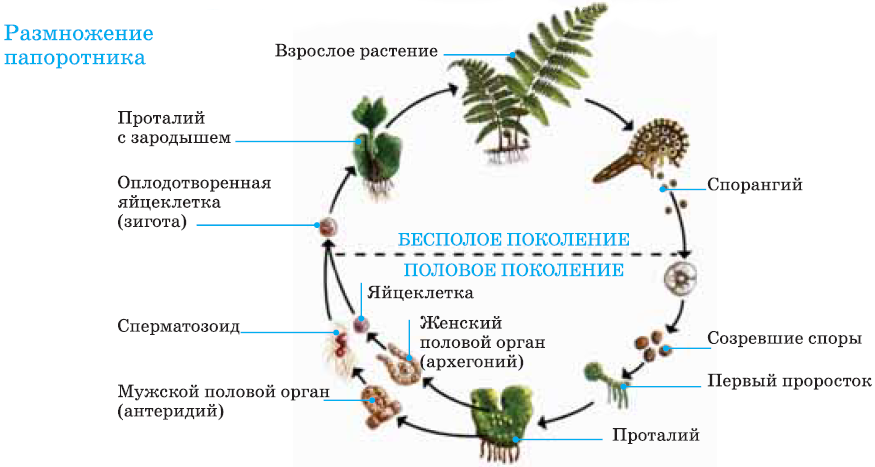
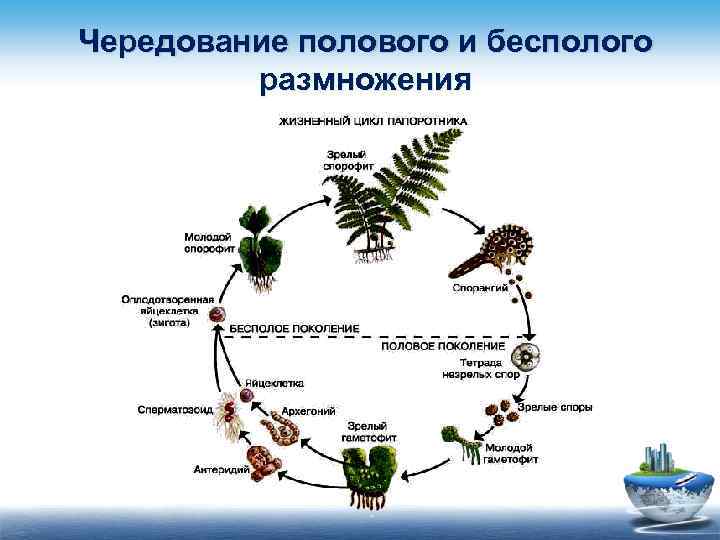
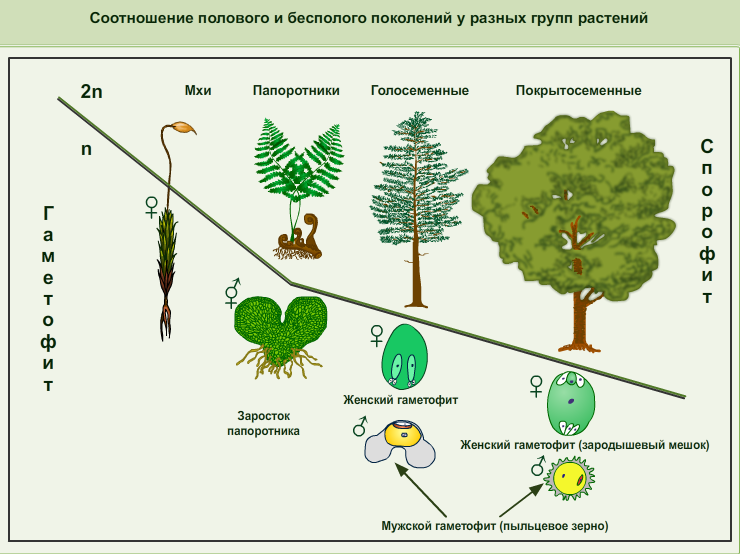
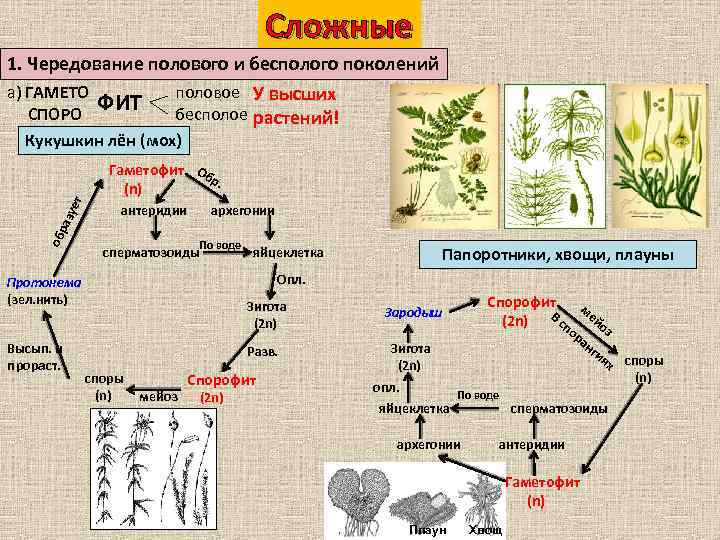
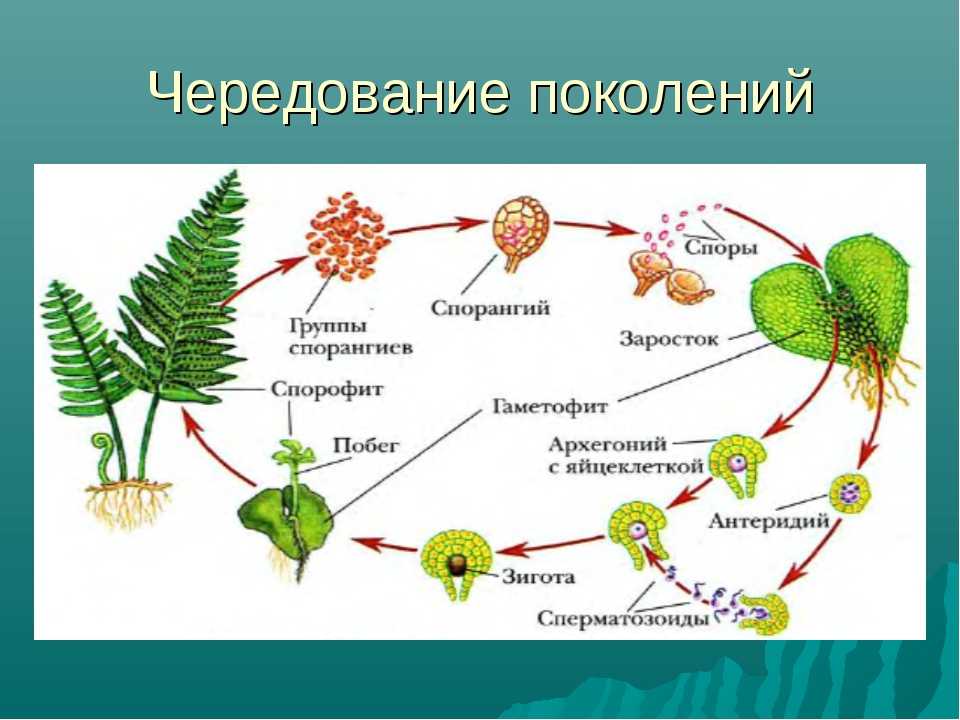
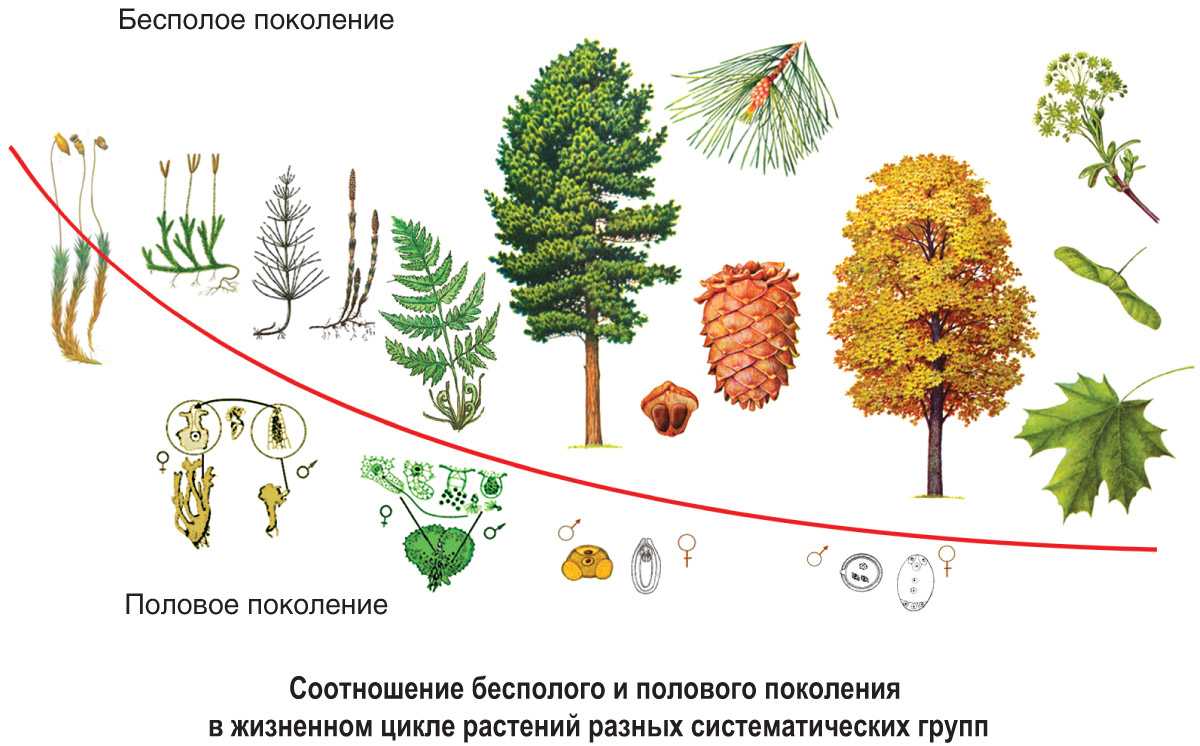


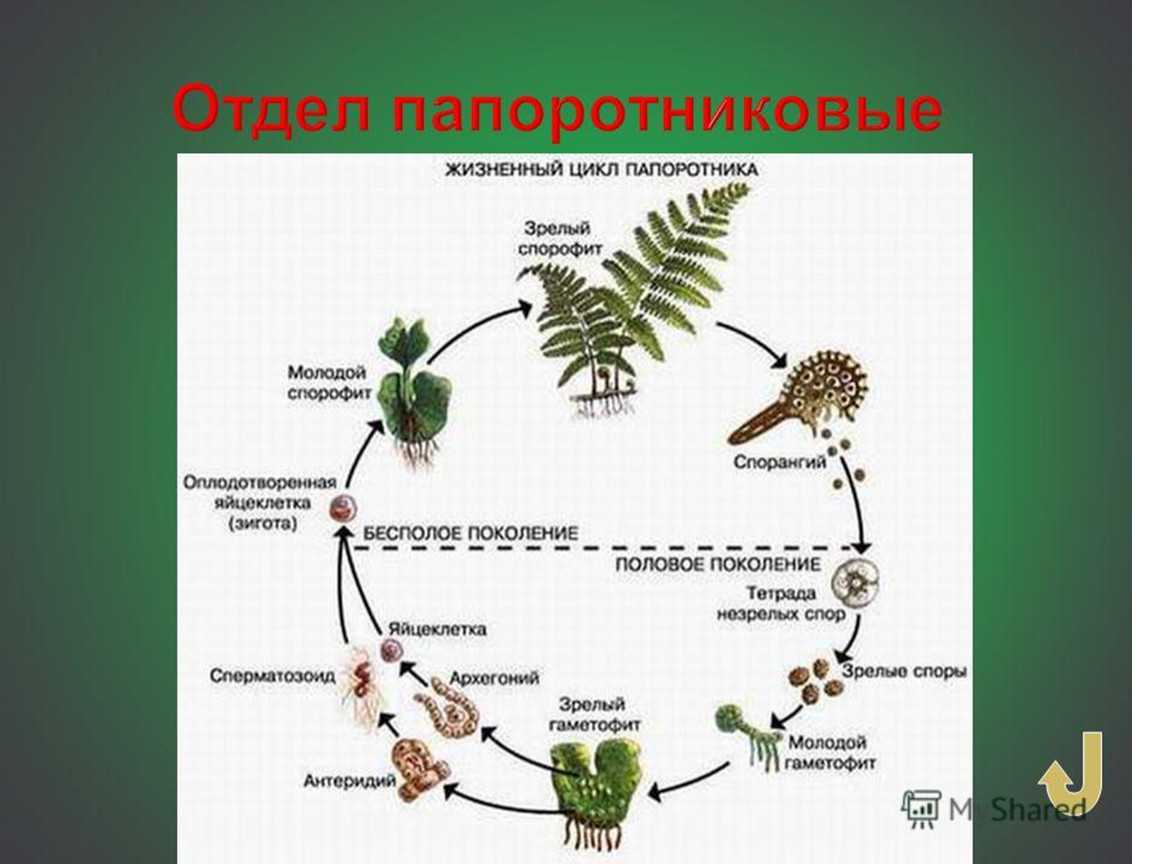
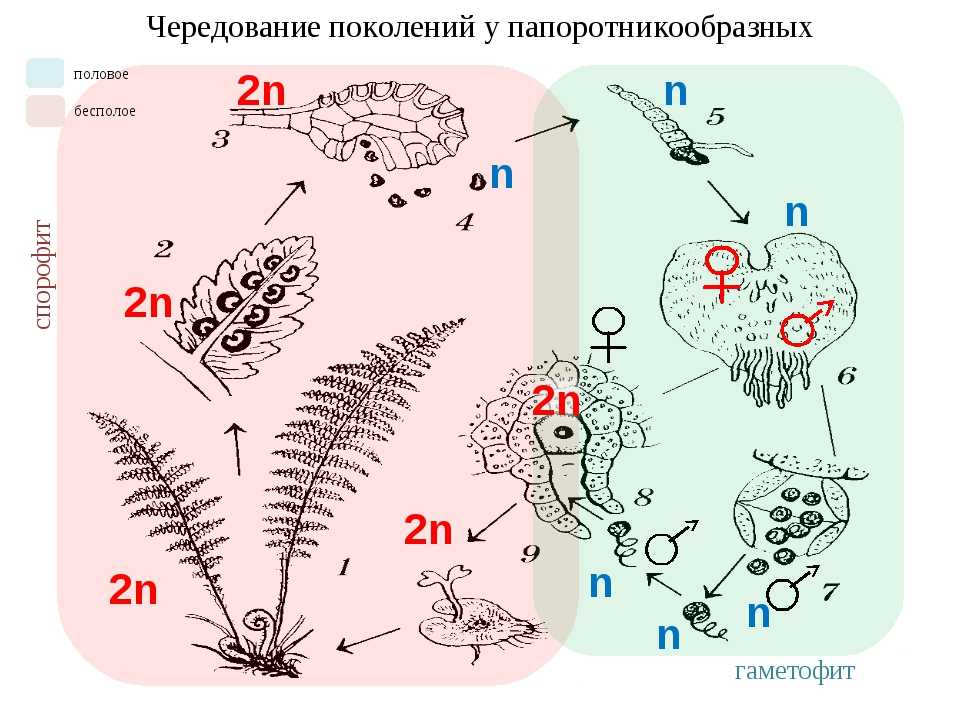
Первичной фазой жизненного цикла сосудистых растений является генерация спорофитов. В сосудистых растениях, которые не производят семена, такие как папоротники и хвощи, поколения спорофитов и гаметофитов независимы. Например, у папоротников ветвь с листьями представляют собой зрелое диплоидное образование спорофитов. Спорангии на нижней стороне листьев вырабатывают гаплоидные споры, которые прорастают для образования гаплоидных гаметофитов папоротника (проталлий). Эти растения процветают во влажных условиях, так как вода необходима для оплодотворения.
Сосудистые растения, которые производят семена, не всегда зависят от влажных сред обитания для размножения. Семена защищают развивающиеся эмбрионы. Как в цветковых, так и в нецветковых растениях (хвойных) генерация гаметофитов полностью зависит от доминирующих поколений спорофит. В цветущих растениях репродуктивная структура — цветок. Цветок производит как мужские микроспоры, так и женские мегаспоры.
Сами микроспоры содержатся в пыльце и вырабатываются в тычинке растения, развиваясь в мужские половые клетки. Женские мегаспоры производятся в пестики растений и развиваются в женские гаметы. Во время опыления пыльца переносится ветром, насекомыми или другими животными в женскую часть цветка. Мужские и женские гаметы объединяются и развиваются в семя, а завязь образует плод. У хвойных, пыльца производится в мужских шишках, а в женских шишках после оплодотворение формируется зародыш.
Мне нравитсяНе нравится1
Половое размножение водорослей
Половое размножение у водорослей связано с половым процессом, который заключается в слиянии двух клеток, в результате чего образуется зигота, вырастающая в новую особь или дающая зооспоры.
Половое размножение у водорослей бывает нескольких типов:
- гологамия (коньюгация) – без образования специализированных клеток;
- гаметогамия – с помощью специализированных клеток – гамет.
Гологамия
В простейшем случае процесс происходит путем слияния двух неподвижных, лишенных клеточных оболочек вегетативных клеток. У одноклеточных жгутиковых форм водорослей половой процесс осуществляется путем слияния двух особей.
При слиянии содержимого двух безжгутиковых вегетативных клеток половой процесс называют коньюгацией.
Во время коньюгации происходит слияние двух клеток, которые выполняют функцию половых клеток – гамет. Слияние содержимого клеток происходит через специально образующийся коньюгационный канал, получается зигота, которая впоследствии покрывается толстой оболочкой и превращается в зигоспору. Если скорость перетекания содержимого клеток одинакова, зигота образуется в коньюгационном канале. В этом случае разделение клеток на мужскую и женскую условно.
Гаметогамия
Половое размножение у водорослей, в том числе и одноклеточных, чаще происходит путем деления содержимого клеток и образования в них специализированных половых клеток – гамет. У всех зеленых и бурых водорослей мужские гаметы имеют жгутики, а женские гаметы имеют их не всегда. У примитивных водорослей гаметы образуются в вегетативных клетках. У более высокоорганизованных форм гаметы находятся в особых клетках, называемых гаметангиями.
В вегетативной клетке или гаметангии может быть от одной до нескольких сотен гамет. В зависимости от размеров сливающихся гамет различают несколько типов гаметогамии: изогамия, гетерогамия, оогамия.
Если сливающиеся гаметы имеют одинаковую форму и размеры, такой половой процесс называют изогамией.
Если сливающиеся гаметы имеют одинаковую форму, но разные размеры (женская гамета крупнее мужской), то говорят о гетерогамии.
Половой процесс, при котором сливаются неподвижная крупная клетка – яйцеклетка и подвижная мелкая мужская клетка – сперматозоид, называется оогамией.
Гаметангии с яйцеклетками называют архегониями или оогониями, а со сперматозоидами – антеридиями. Мужские и женские гаметы могут развиваться на одной особи (однодомные) или на разных особях (двудомные).
Образующаяся в результате слияния гамет зигота после некоторых изменений превращается в зигоспору. Последняя обычно одета плотной оболочкой. Зигоспора может находиться длительное время в покое (до нескольких месяцев) или прорастать без периода покоя.
Автогамия. Особый тип полового процесса. Он заключается в том, что ядро клетки делится мейотически, из четырех образовавшихся ядер, два разрушаются, а оставшиеся два ядра сливаются, образуя зиготу, которая без периода покоя увеличивается в размерах и превращается в ауксоспору.
Так происходит омоложение особей.
Большое значение полового размножения для эволюции состоит в том, что
Vlad ustelyomov
Половое размножение – более прогрессивная форма размножения, очень широко распространено в природе, как среди растений, так и среди животных. Образующиеся в процессе полового размножения организмы отличаются друг от друга генетически, а также по характеру приспособленности к условиям обитания.
При половом размножении материнским и отцовским организмами вырабатываются специализированные половые клетки – гаметы. Женские неподвижные гаметы называются яйцеклетками, мужские неподвижные – спермиями, а подвижные – сперматозоидами. Эти половые клетки сливаются с образованием зиготы, т. е. происходит оплодотворение. Половые клетки, как правило, имеют половинный набор хромосом (гаплоидный) , так что при их слиянии восстанавливается двойной (диплоидный) набор, из зиготы развивается новая особь. При половом размножении потомство образуется при слиянии гаплоидных ядер. Гаплоидные ядра образуются в результате мейотического деления.
Мейоз ведет к уменьшению генетического материала вдвое, благодаря чему количество генетического материала у особей данного вида в ряду поколений остается постоянным. Во время мейоза происходит несколько важных процессов: случайное расхождение хромосом (независимое расчленение) , обмен генетическим материалом между гомологичными хромосомами (кроссинговер) . В результате этих процессов возникают новые комбинации генов. Поскольку ядро зиготы после оплодотворения содержит генетический материал двух родительских особей, это повышает генетическое разнообразие внутри вида. Если суть и биологическое значение полового процесса едины для всех организмов, то его формы очень разнообразны и зависят от уровня эволюционного развития, среды обитания, образа жизни и некоторых других особенностей.
Половое размножение имеет очень большие эволюционные преимущества по сравнению с бесполым. Сущность полового размножения заключается в объединении в наследственном материале потомка генетической информации из двух разных источников – родителей. Оплодотворение у животных может быть наружным или внутренним. При слиянии образуется зигота с двойным набором хромосом.
В ядре зиготы все хромосомы становятся парными: в каждой паре одна из хромосом отцовская, другая – материнская. Дочерний организм, который разовьется из такой зиготы, в одинаковой мере снабжен наследственной информацией обоих родителей.
Биологический смысл полового размножения состоит в том, что возникающие организмы могут сочетать полезные признаки отца и матери. Такие организмы более жизнеспособны. Половое размножение играет важную роль в эволюции организмов.
§ 30. Понятие полового размножения и полового процесса
Понятие полового размножения и полового процесса
Как вы уже знаете из § 29—1, половое размножение протекает с участием половых клеток (гамет). Оно обеспечивает наследственное разнообразие потомства и повышает его приспособленность к условиям среды обитания.
Половой процесс — биологическое явление, приводящее к обмену наследственным материалом между особями одного вида или к его объединению, что создает условия для возникновения разнообразия наследственной информации.
Половой процесс представляет собой начало полового размножения, происходящего с участием гамет. Но эти два явления нельзя отождествлять, так как половой процесс не всегда приводит к увеличению числа особей. Иногда половой процесс отождествляют с оплодотворением, т. е. слиянием женской и мужской гамет. Этого тоже делать нельзя, поскольку половой процесс может протекать и без участия гамет (водоросли, инфузории).
Формами полового процесса являются конъюгация и копуляция.
Конъюгация—особая форма полового процесса, при которой происходит контакт одноклеточных организмов или соматических клеток многоклеточных организмов с образованием цитоплазматических мостиков для перехода ядер или всего содержимого клеток. Конъюгация у одноклеточных организмов (инфузории) несколько отличается от таковой у многоклеточных водорослей (спирогира). У инфузорий (например, инфузории туфельки) во время коньюгации две особи вступают в контакт с помощью цитоплазматического мостика, через который обмениваются подвижными малыми ядрами. Большие ядра у них погибают и в половом процессе не участвуют. При этом не происходит увеличения числа особей, но обеспечивается перекомбинация наследственного материала.
При конъюгации у многоклеточных водорослей (спирогира) отдельные гаплоидные клетки параллельно лежащих нитевидных талломов попарно образуют навстречу друг другу выросты, которые соединяются и формируют цитоплазматический мостик. Через этот мостик содержимое одной из клеток перетекает в другую и соединяется с ее содержимым. При этом происходит направленный перенос наследственной информации из одной вегетативной клетки в другую, которая превращается в диплоидную клетку. Последняя делится на две дочерние клетки, дающие начало двум новым гаплоидным особям. При этом количество особей будет увеличиваться, так как в конъюгации могут участвовать многие клетки многоклеточных нитей водоросли.
Копуляция (гаметогамия) — форма полового процесса, при которой у многоклеточных и одноклеточных организмов две различающиеся по полу клетки (гаметы) сливаются и образуют зиготу, содержащую одно ядро с новым набором наследственного материала. Из зиготы развивается новый организм.
*Различают следующие основные формы гаметогамии:
- изогамия — гаметы не отличаются друг от друга по размерам, подвижны, имеют жгутики, однако физиологически они разделяются на «мужскую» и «женскую». Изогамия встречается у некоторых протистов и многих водорослей;
- анизогамия (гетерогамия) — гаметы отличаются друг от друга морфологически (одна крупнее другой) и физиологически, но оба типа гамет подвижны и имеют жгутики. Такой тип полового процесса характерен для некоторых водорослей;
- оогамия (овогамия) — в данном случае гаметы сильно отличаются друг от друга и разделяются на женские — яйцеклетки и мужские — сперматозоиды. Яйцеклетки крупные и неподвижные, а сперматозоиды мелкие и подвижные. У семенных растений мужские гаметы неподвижны и называются спермиями. Они доставляются к яйцеклетке с помощью пыльцевой трубки. Оогамия характерна для животных, растений и многих грибов.
Копуляция как форма полового процесса эволюционировала в следующем направлении:
изогамия → анизогамия → оогамия.
Эмбриональное развитие гипоталамо-гипофизарно-яичниковой оси и женских половых органов
Развитие ГГЯ оси и женских половых органов — сложный и ответственный процесс, определяющий репродуктивные возможности женщины.
Эмбриология нейронов, секретирующих гонадолиберин. Нейроны, секретирующие гонадолиберин, поступают из клеток-предшественников из обонятельного эпителия. Мигрируя вместе с обонятельными нейронами, они проходят через решетчатую кость в передний мозг. Обонятельные нейроны образуют обонятельную луковицу, а нейроны, выделяющие ГнРГ, расположены в основном в дугообразном ядре и в поле предварительного зрения.

За успешный ход миграции и развития нейронов отвечают адгезивные молекулы, регулирующие движение и удлинение нейронов. Миграция заканчивается примерно на 8-й неделе беременности.
Эмбриология яичников. В раннем периоде эмбрионального развития промежуточная мезодерма образует мочеполовую складку, из которой развиваются гонады, мюллеровские и вольфовские протоки.
Первичные половые клетки появляются в эмбриональном развитии уже около 3-х недель, в энтодерме фолликула желтка. Между 4 и 5 неделями 300-1000 клеток начинают мигрировать через брыжейку к половым складкам и становятся оогониями — незрелыми половыми клетками, способными к митозу. Органы выделения и половые протоки образуются из промежуточной мезодермы.
В течение этого времени в оогонии происходит процесс генетического импринтинга — метилирование CpG островков. Метилирование препятствует прикреплению белков транскрипционного аппарата. Результат процесса — отсутствие экспрессии одной из двух аллелей (формы одного гена) материнских или отцовских генов в клетках потомства. Изменения, происходящие в клетке, определяют здоровье будущего потомства.
В этот период гонада морфологически недифференцирована, но наличие половых клеток индуцирует ее развитие по хромосомному полу. В недифференцированных половых железах половые клетки интенсивно размножаются до 16-20 недель. В гонаде плода на 20 неделе беременности количество половых клеток достигает 7 млн.
На 11-12 неделе гестации начинается первое редукционное деление ДНК в оогонии, но останавливается на профазной стадии до овуляции у взрослой женщины. Клетки в позднем диплотене окружены клетками, полученными из половых тяжей, и начинается образование фолликулов. В яичнике плода постоянно прогрессирует рост фолликулов до преантральной фазы и атрезия оогонии и фолликулов. Апоптоз (запрограммированный процесс гибели клеток) практически устраняет все оогонии к моменту родов, а количество фолликулов уменьшается до 2 млн.
Эмбриология матки, маточных труб и влагалища. Матка развивается из двух мюллеровых протоков, появляющихся у обоих полов на 6-7 неделе эмбрионального развития.
- Плодная мужская гонада вырабатывает антимюллеров гормон (АМГ), вызывающий лизис (растворение клеток) протоков Мюллера.
- Женская гонада начинает синтез АМГ только в период полового созревания.
У плодов женского пола протоки Мюллера развиваются и образуют фаллопиевы трубы, матку и верхнюю часть влагалища. В промежуточной части протоков примерно на 9-й неделе беременности протоки сближаются и сливаются, образуя маточно-влагалищный канал.
Что такое животный секс?
Основное понятие животного секса заключается в том, что животные используют свои половые органы для размножения и передачи генетической информации. У разных видов животных сексуальное поведение может проявляться по-разному.
Животный секс имеет свои особенности, отличающие его от секса у людей. В отличие от людей, у животных может отсутствовать эмоциональная составляющая в сексуальных отношениях. Животные часто имеют инстинкты, которые управляют их сексуальным поведением и заставляют их размножаться.
Интересно отметить, что животный секс включает в себя разнообразные стратегии и приемы, которые помогают животным достичь успешного размножения. Некоторые виды животных имеют сложные ритуалы и игры перед сексуальным актом, другие используют разнообразные механизмы, такие как самооплодотворение или пути размножения без полового контакта.
Таким образом, понятие животного секса включает в себя различные формы сексуальной активности животных, которые направлены на размножение и передачу генетической информации. Это уникальный аспект жизни животных, который помогает им выживать и сохранять виды.
Определение животного секса
Животный секс включает различные аспекты, такие как спаривание, оплодотворение, беременность и роды. Процесс сексуального взаимодействия может быть разнообразным и иногда сложным в зависимости от вида животного.
Определение животного секса помогает понять эволюцию и разнообразие размножения в животном мире. Изучение животного секса позволяет выявить особенности размножения разных видов животных и понять, как эти особенности влияют на их поведение и биологическую адаптацию к окружающей среде.
Функции и цели животного секса
Основные функции животного секса:
- Размножение: Одной из главных функций животного секса является возможность размножения и производства потомства для продолжения видов. Сексуальное размножение позволяет создавать генетически разнообразные потомки, что повышает их шансы на выживание, а также способствует приспособлению и эволюции видов.
- Генетическая разнообразность: Сексуальное размножение способствует генетической разнообразности в популяции. Комбинирование генетического материала от двух родителей позволяет создавать потомство с новыми комбинациями генов, что способствует адаптации к изменяющимся условиям окружающей среды.
- Укрепление социальных связей: У некоторых видов животных, сексуальная активность может способствовать укреплению социальных связей и способствовать образованию и поддержанию парных связей между особями. Например, у некоторых птиц и млекопитающих проявление сексуальности может быть связано с созданием парных бондов, заботой о потомстве и сотрудничеством при поиске пищи и защите от хищников.
- Наследование ритуалов и поведенческих особенностей: Сексуальный процесс может быть связан с наследованием ритуалов и поведенческих особенностей от предков. Различные виды животных проявляют определенные ритуалы и поведения во время сексуального акта, которые передаются от поколения к поколению и служат сигналами и символами для привлечения партнера.
Животный секс имеет свои особенности в зависимости от вида и видового поведения. Отсутствие сексуальности у некоторых видов животных может быть связано с различными факторами, такими как размножение путем деления клеток или самооплодотворение.
Клиническая оценка функции яичников — овариальный резерв
Репродуктивный потенциал яичников (биологический возраст) называют овариальным резервом. Этот термин описывает количество антральных фолликулов, готовых реагировать на экзогенные гонадотропины или пул фолликулов на антральной стадии в яичнике.
Овариальный резерв генетически детерминирован и снижается по следующим причинам:
- с возрастом;
- в результате операций на яичнике;
- в результате патологий яичников.
Оценка овариального резерва позволяет спрогнозировать реакцию яичника на гонадотропную стимуляцию, определить метод стимуляции и дает возможность понять, насколько вероятна беременность. Прогрессирующее снижение овариального резерва с возрастом сопровождается снижением качества яйцеклеток, выраженным, в том числе, увеличением хромосомных нарушений.
Для клинической оценки овариального резерва применяются следующие методы диагностики:
| Параметр | Показатель | Выводы |
| Количество антральных фолликулов размером 2-4 мм | <5 | предсказывает плохую реакцию на гонадотропины. |
| Объем яичников | <5 см | низкая вероятность ответа на гонадотропины. |
| Концентрация ФСГ на 3-й день цикла | >10 МЕ/л | снижение овариального резерва. |
| >20 МЕ/л | истечение репродуктивной функции. | |
| Концентрации эстрадиола на 3-й день цикла (низкая полезность) | >80 пг/мл вместе с повышенной концентрацией ФСГ | быстрое выделение доминантного фолликула из небольшого пула фолликулов яичников. |
| Концентрация ингибина В (низкая полезность) | >45 пг/мл | плохая реакция на стимуляцию. |
| Концентрация АМГ | >20 пмоль/л | чрезмерная реакция яичников на гонадотропную стимуляцию. |
| <1 пмоль/л | ответ на стимуляцию маловероятен. |
Оценка качества полового цикла
Диагностика качества цикла заключается в подтверждении произошедшей овуляции и других необходимых для беременности явлений.
Для этого используются следующие методы диагностики:
- Оценка цервикальной слизи. В фазе роста фолликула слизь скудная, липкая. В периовуляторном периоде слизь обильная, водянистая, прозрачная. В лютеиновой — снова скудная, мутная, липкая.
- Измерение базальной температуры тела. В фолликулярной фазе температура тела колеблется от 36,1 до 36,6°С, а в лютеиновой увеличивается на 0,4-0,8°С. Причина повышения температуры — термогенное действие метаболитов прогестерона. Самая высокая фертильность наблюдается за 7 дней до повышения температуры тела.
- Измерение концентрации прогестерона за 7 дней до ожидаемой менструации. Концентрация >5 нг/мл свидетельствует об овуляции. Но нужно понимать, что на основании результатов анализа на концентрацию прогестерона нельзя оценить эффективность желтого тела.
- Фолликулометрия и УЗИ матки. УЗИ размеров фолликула Граафа и толщины эндометрия, проводят в комплексе с оценкой концентрации эстрадиола. В периовуляторном периоде фолликул достигает 16-20 мм, а концентрация эстрадиола — 150-350 пг/мл.
Функция матки в менструальном цикле
В эндометриальном слое матки происходят процессы, позволяющие имплантироваться и развиваться плодному яйцу. Эндометрий состоит из функционального и основного слоев и является уникальной тканью, поскольку его морфология периодически изменяется в менструальном цикле:
- При менструации эндометрий отслаивается до толщины <2 мм;
- При повышении концентрации эстрадиола, продуцируемого растущим фолликулом Граафа, начинается пролиферативная фаза роста эндометрия. Повреждённые ткани восстанавливаются. В ранней фазе железы узкие, прямые, трубчатые, клетки кубические, с круглым ядром. В поздней – из-за гиперплазии желез и накопления межклеточного матрикса в строме эндометрий становится толстым, железистые клетки перекрываются.
- После овуляции в эндометрии начинается фаза секреции.
Эта фаза также делится на этапы с характерной морфологической картиной:
- В первые сутки железы сферические.
- На второй день они становятся более овальными в поперечном сечении, присутствуют митозы, а гидроцеле встречается менее чем в половине желез.
- На четвертый день митоз отсутствует, железистый эпителий высокий.
- На шестой день обнаруживаются выделения в железах, железы нерегулярно расширяются, в строме имеются голые клеточные ядра.
- На седьмой день наблюдается отек стромы, в просвете желез еще есть секреция, железистые клетки становятся кубовидными.
- На 9-й день после овуляции начинается псевдодецидуализация.
Морфология эндометрия может помочь при диагностике нарушений менструального цикла, но сегодня биопсия эндометрия с этой целью не назначается.
Размножение водорослей
Каким образом размножаются водоросли?
Водоросли размножаются половым и бесполым путем.
Как правило, бесполым путем водоросли размножаются в благоприятный период.
Бесполое размножение осуществляется вегетативно (у одноклеточных — делением надвое, у многоклеточных — частями слоевища), спорами (неподвижными клетками) и зооспорами (подвижными клетками). Рассмотрим для примера бесполое (зооспорами) и половое размножение одноклеточной водоросли. Если водоросль подвижна, то перед размножением она теряет жгутики. Ядро и цитоплазма делятся пополам; затем происходит еще одно или два деления, в результате которых в одной и той же оболочке образуются 4–8 клеток.

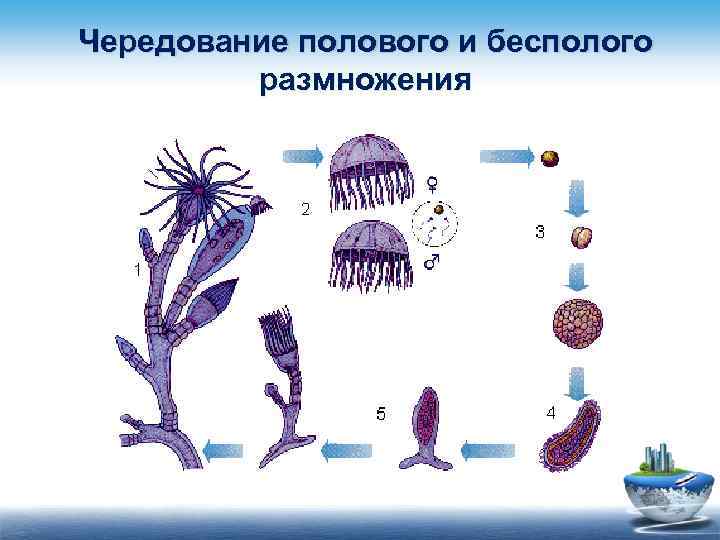

Эти мелкие подвижные клетки — зооспоры — выходят из оболочки материнской клетки и вырастают во взрослую особь.
В неблагоприятный период (высокая или низкая температура, накопление продуктов обмена в среде обитания при высокой плотности заселения, загрязнение водоемов) происходит половое размножение. Каждая клетка делится на много мелких половых клеток — гамет, которые потом попарно сливаются с гаметами другой особи, образуя зиготу, несущую в результате признаки обеих родительских особей.
Последняя покрывается плотной оболочкой и зимует. Весной из зиготы выходит 4 зооспоры. Каждая из них дает начало новому растению.
Изогамный половой процесс
При изогамии (равный брак) гаметы морфологически сходны между собой, то есть одинаковы по форме и размерам, но физиологически они разнокачественны.
Данный половой процесс характерен для многих водорослей и некоторых грибов. Изогамия происходит только в воде, для передвижения в которой гаметы снабжены жгутиками. Они очень похожи на зооспоры, но имеют меньшие размеры.
При гетерогамии (разный брак) происходит слияние подвижных половых клеток, сходных по форме, но различающихся размерами. Женская гамета в несколько раз больше мужской и менее подвижна. Гетерогамия характерна для тех же групп организмов, что и изогамия, и также происходит в воде.
Характерна для некоторых водорослей и всех высших растений. Женская гамета — яйцеклетка — крупная и неподвижная. У низших растений образуется в одноклеточных гаметангиях — оогониях, у высших растений (исключая покрытосеменные) — в многоклеточных архегониях.
Мужская гамета (сперматозоид) мала и подвижна, образуется у грибов и водорослей в одноклеточных, а у высших растений (исключая покрытосеменные) — в многоклеточных гаметангиях — антеридиях.
Сперматозоиды способны передвигаться только в воде. Поэтому наличие воды — обязательное условие для оплодотворения у всех растений, за исключением семенных. У большинства семенных растений мужские гаметы утратили жгутики и называются спермиями.