
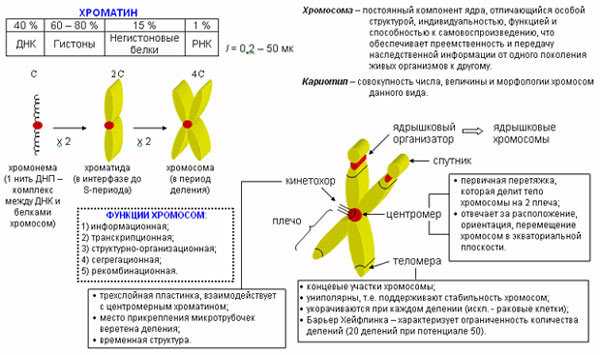
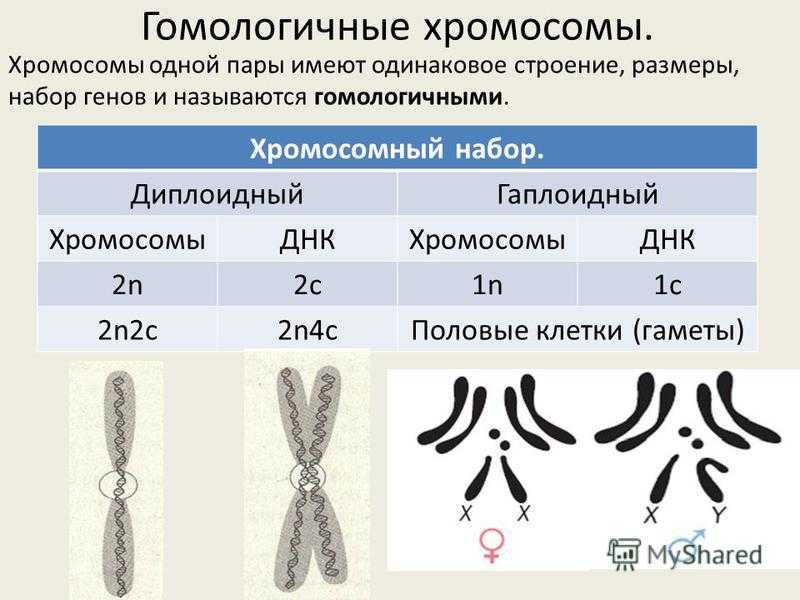
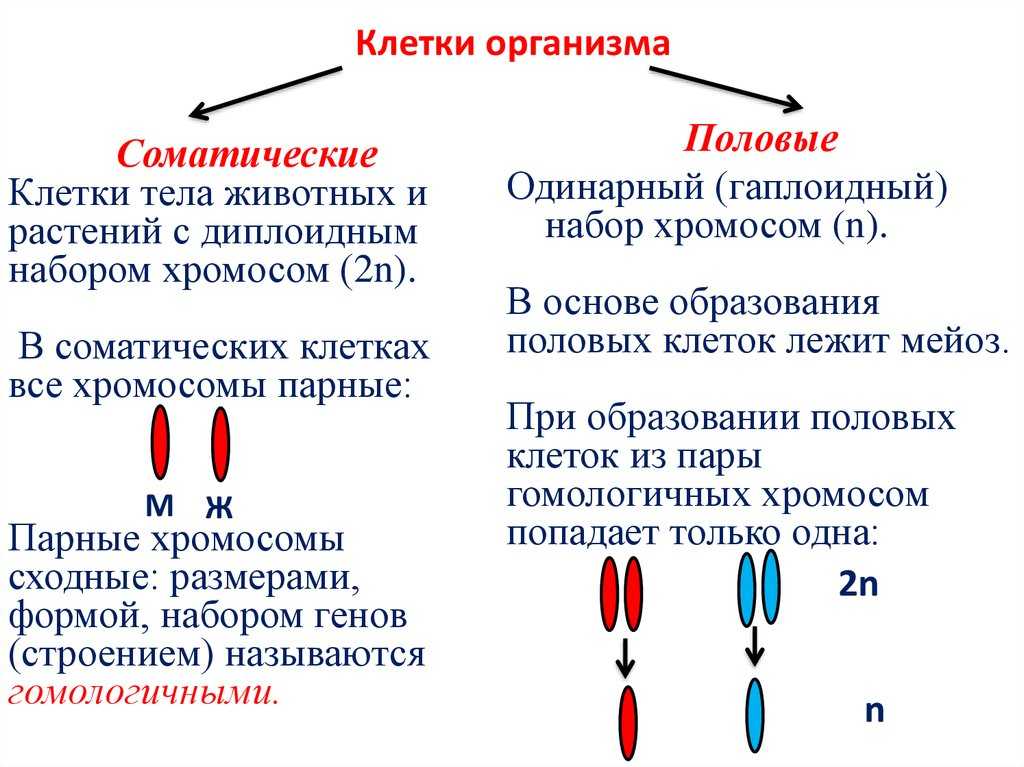
Содержание
А дупликация хромосомы описывает фракцию ДНК, которая дважды появляется как продукт генетической рекомбинации. Хромосомная дупликация, дупликация или амплификация генов — один из источников генерации изменчивости и эволюции живых существ.
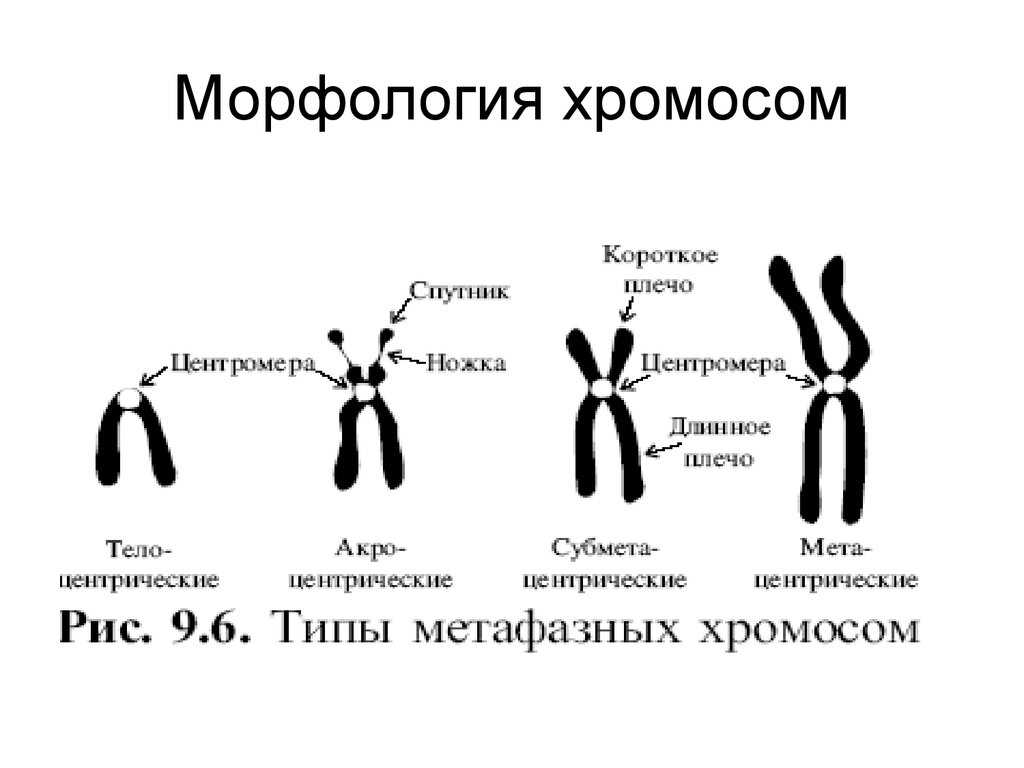
Хромосомная дупликация — это тип мутации, поскольку она включает изменение нормальной последовательности ДНК в хромосомной области. Другие мутации на хромосомном уровне включают хромосомные вставки, инверсии, транслокации и делеции.
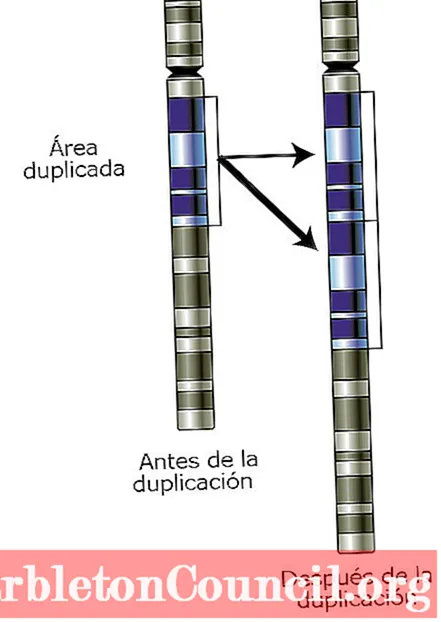
Хромосомные дупликации могут происходить на том же исходном сайте, что и дублированный фрагмент. Это пакетные дубликаты. Пакетные дубликаты могут быть двух типов: прямые и инвертированные.
Прямые дубликаты — это те, которые повторяют как информацию, так и ориентацию повторяющегося фрагмента. В пакетно инвертированных повторяющихся фрагментах информация повторяется, но фрагменты ориентированы в противоположных направлениях.
В других случаях хромосомная дупликация может происходить на другом участке или даже на другой хромосоме. Это генерирует эктопическую копию последовательности, которая может функционировать как субстрат для кроссовера и быть источником аберрантных рекомбинаций. В зависимости от размера дубликаты могут быть макро- или микродупликациями.
С эволюционной точки зрения дублирование порождает изменчивость и изменения. Однако на индивидуальном уровне дупликация хромосом может привести к серьезным проблемам со здоровьем.
Дублирование гена
Механизмы дупликации генов
Конкретный ген может дублироваться в процессе эволюции. Существует четыре основных механизма, позволяющих дублировать один или несколько генов:
- Путем ретротранспозиции : во время ретротранспозиции мобильного элемента может происходить транскрипция части хромосомной области, близкой к мобильному элементу, необязательно содержащей один или несколько генов. Когда РНК ретранскрибируется в геноме, случайно транскрибированные ген или гены копируются в другую область генома.
- Путем неравного кроссинговера (во время мейоза гомологичные хромосомы объединяются в пары на одинаковых участках и в конечном итоге могут обмениваться). Таким образом, часть ДНК одной из двух гомологичных хромосом переносится на другую. Таким образом, происходит потеря генов на одной хромосоме и дупликация генов на другой, если эта часть хромосомы несет один или несколько генов. Затем мы получаем расположение генов, продублированных один за другим в тандеме.
- Путем эктопического обмена : во время двухцепочечного разрыва может происходить рекомбинация с негомологичным сайтом, затем удлинение молекулы ДНК по матрице рекомбинантной ДНК, таким образом копируя часть генома после разрыва. Затем восстановление разрыва приводит к интеграции этой дублированной области в геном.
- Путем горизонтального переноса генов механизма : Экзогенный ДНК происходит из организма преобразователя ( вирус , паразит , мертвые клетки, эндосимбионта ( митохондрии , хлоропласт )). Однако этот механизм обмена генетическим материалом редко вызывает дупликацию генов.
Что такое репликация
Репликация — это процесс, который синтезирует идентичную копию двухцепочечных молекул ДНК в клетке. Это происходит как в эукариотических, так и в прокариотических клетках. Основная функция репликации ДНК заключается в увеличении количества ДНК в клетке в два раза, поскольку клетка подвергается делению с образованием дочерних клеток. Это позволяет каждой дочерней клетке содержать точное количество ДНК в качестве родительской клетки.
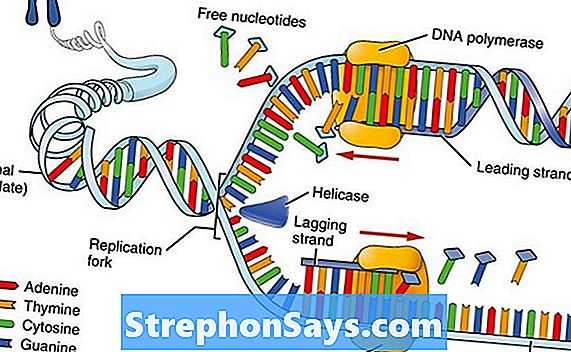
Рисунок 1: Репликация ДНК
Как правило, ДНК представляет собой двухцепочечную молекулу, состоящую из двух цепей ДНК. Каждая цепь ДНК служит шаблоном для синтеза новой ДНК. Основным ферментом, ответственным за репликацию ДНК, является ДНК-полимераза. Кроме того, в этом процессе участвуют несколько ферментов, в том числе ДНК-геликаза, РНК-примаза и т. Д. ДНК-геликаза раскручивает двухцепочечную ДНК, образуя репликационную вилку. Затем РНК-примаза добавляет праймер к шаблонам ДНК, чтобы инициировать репликацию ДНК. Затем ДНК-полимераза добавляет комплементарные нуклеотиды к матричной цепи. Поступающие нуклеотиды ковалентно связаны с вновь синтезированной цепью ДНК через фосфодиэфирные связи. Репликация ДНК происходит полуконсервативным образом, поскольку одна цепь каждой вновь синтезированной двухцепочечной ДНК является старой цепью.
ссылки
- Cordovez, J.A., Capasso, J., Lingao, M.D., Sadagopan, K.A., Spaeth, G.L., Wasserman, B.N., Levin, A.V. (2014). Глазные проявления микродупликации 22q11.2. Офтальмология, 121: 392-398.
- Goodenough, U. W. (1984) Genetics. W. B. Saunders Co. Ltd, Филадельфия, Пенсильвания, США.
- Griffiths A.J.F., Wessler R., Carroll S.B., Doebley J. (2015). Введение в генетический анализ (11-е изд.). Нью-Йорк: У. Х. Фриман, Нью-Йорк, Нью-Йорк, США.
- Hardison, R.C. (2012) Эволюция гемоглобина и его генов. Перспективы гавани Колд Спринг в медицине 12, дои: 10.1101 / cshperspect.a011627
- Вайс, А., Мрасек, К., Кляйн, Э., Мулатино, М., Ллерена-младший, Ю.С., Хардекопф, Д., Пекова, С., Бхатт, С., Косякова, Н., Лиер, Т. (2012) Синдромы микроделеции и микродупликации. Журнал гистохимии и цитохимии 60, doi: 10.1369 / 0022155412440001
Дупликация простыми словами для чайников
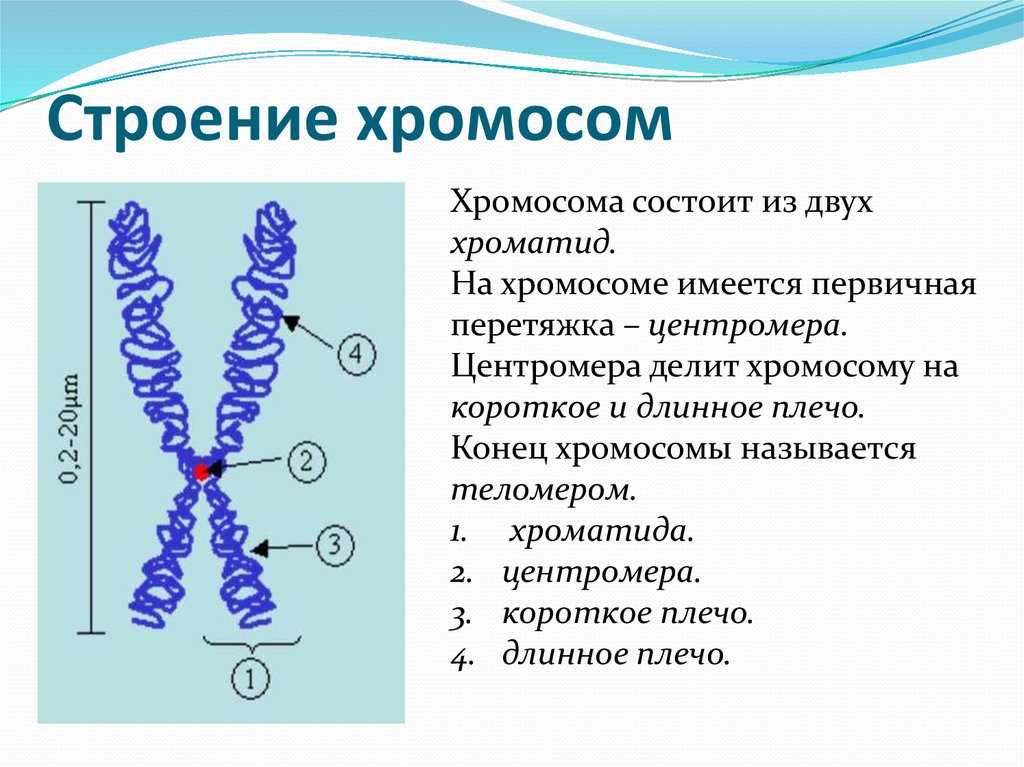
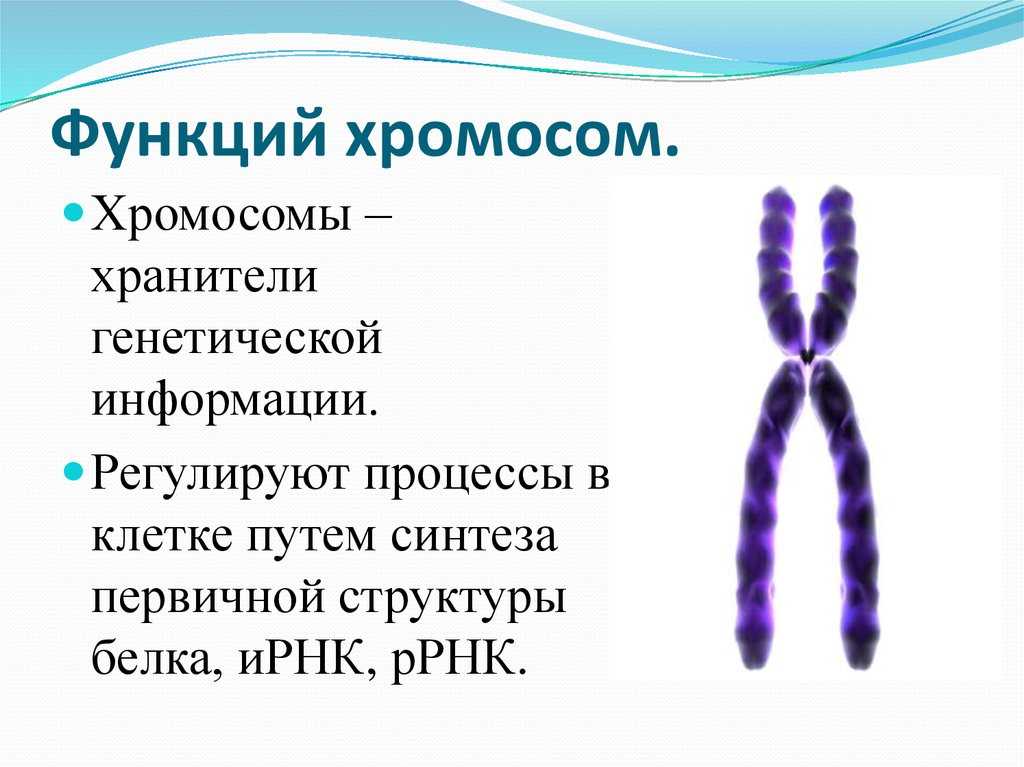
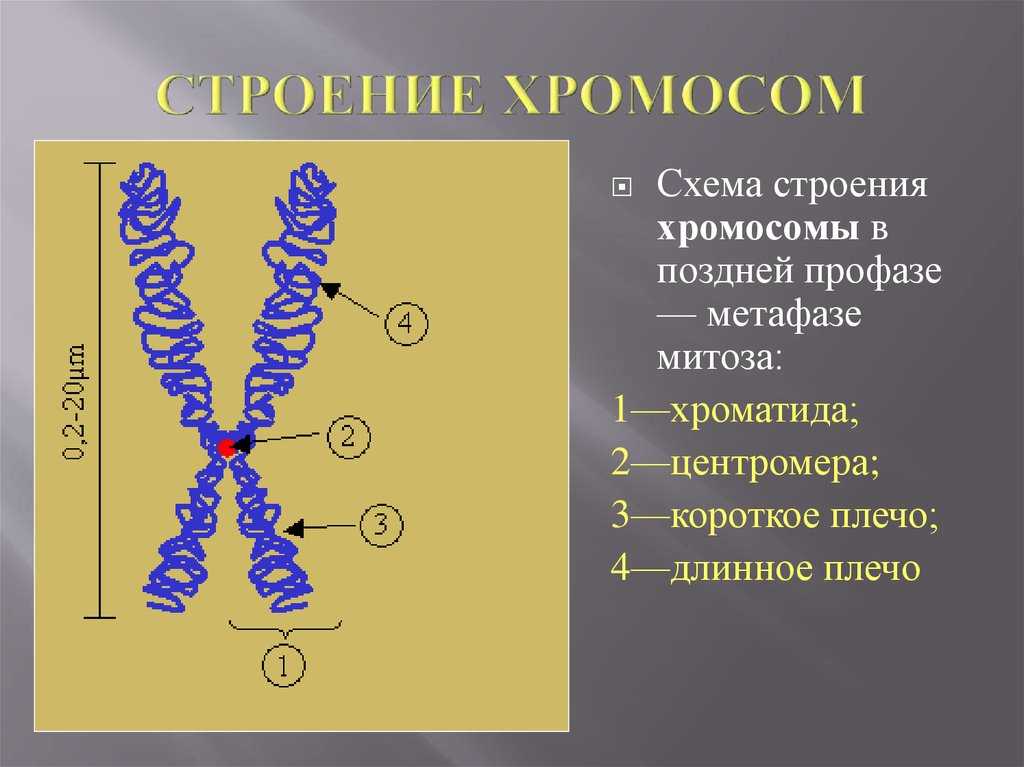
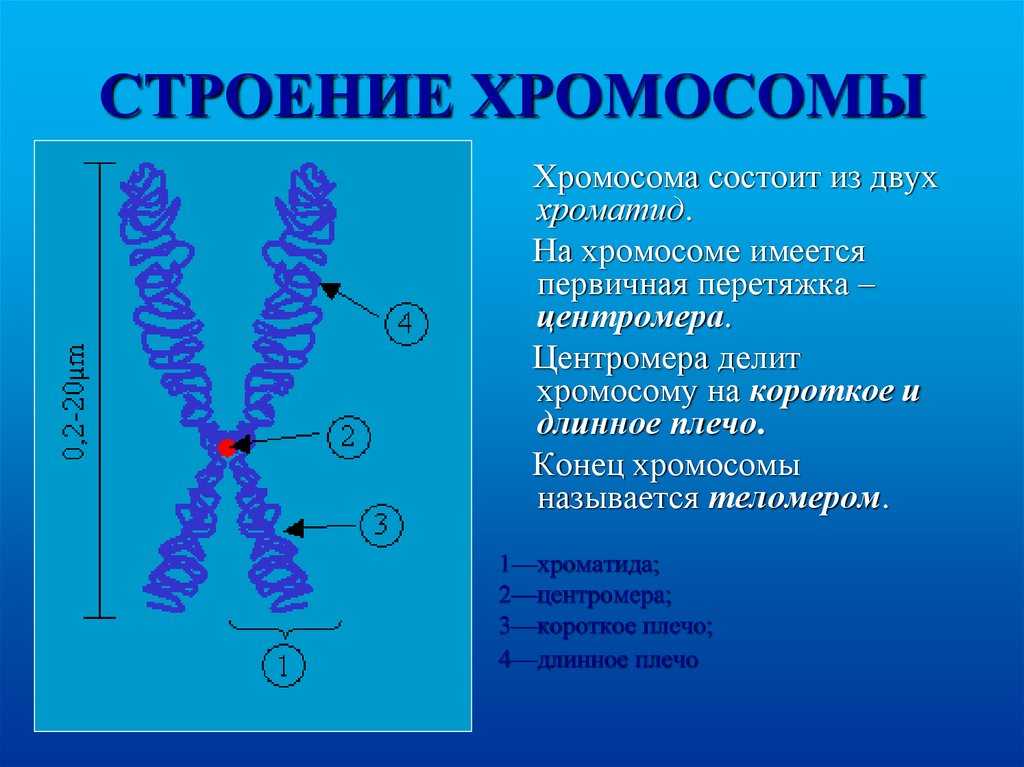
Дупликация — это генетическое изменение, которое происходит на уровне хромосом, нитевидных структур, содержащих нашу генетическую информацию. В этом случае происходит повторение какого-либо участка хромосомы.
Чтобы лучше понять, представьте, что хромосома — это как книга с генетической информацией, а гены — это слова или предложения в этой книге. Дупликация подобна ситуации, когда некоторые страницы или абзацы в книге повторяются.
В результате дупликации, участок хромосомы, содержащий определенные гены, повторяется один или несколько раз. Это может произойти из-за ошибки при делении хромосом в процессе размножения клеток или из-за воздействия некоторых факторов окружающей среды.
Дупликации могут быть как видимыми, то есть приводить к явным изменениям в организме, так и невидимыми, не проявляющимися сразу. Видимые дупликации могут вызывать различные заболевания и синдромы, так как повторение генов может привести к неправильной работе организма.
Например, дупликация гена, отвечающего за рост клеток, может привести к неправильному развитию органов, что может вызвать различные патологии. Также дупликация генов, связанных с иммунной системой, может привести к нарушениям в работе иммунной системы и повышенной восприимчивости к инфекционным заболеваниям.
Однако не все дупликации являются вредными. Иногда они могут быть полезными и даже способствовать эволюции организма. Дупликации генов могут создавать дополнительные копии генетической информации, что дает организму больше возможностей для изменений и адаптации к изменяющейся среде.
В целом, дупликация — это одна из мутаций, которые могут происходить на генетическом уровне. Она может вызывать различные изменения в организме, как положительные, так и отрицательные. Однако, чтобы полностью понять все последствия дупликации, требуется дальнейшее исследование и изучение этого процесса.

Дупликация гена
Дупликация генов была первым предложенным механизмом генерации генов (Ohno, 1970), и этот процесс действительно кажется наиболее распространенным способом создания новых генов. Дупликации обычно классифицируются по размеру дублируемой части генома ; таким образом, дупликация может быть описана как затрагивающая весь геном, большие сегменты генома, отдельные гены, отдельные экзоны или даже определенные части экзонов (Betrán & Long, 2002). Механизмы, которые генерируют повторяющиеся гены, разнообразны, и постоянно открываются новые подробности об этих механизмах. Эти механизмы включают дупликации всего генома, происходящие из-за нерасхождения., тандемные дупликации, возникающие в результате неравного кроссовера, ретропозиции, возникающие в результате ретротранскрипции промежуточного соединения РНК, транспозиции с участием мобильных элементов (Jiang et al ., 2004; Morgante et al ., 2005) и дупликации, возникающие после перегруппировок и последующей репарации смещенных разрывов ( Ранц и др ., 2007). В таких дупликациях участвуют не только гены, кодирующие белки, но и гены некодирующих РНК . Напр., Новый класс ретродукатов включает snoRNAs, которые представляют собой класс генов РНК, которые участвуют в процессинге рибосомной РНК (Weber, 2006).
Большая часть нынешнего ажиотажа по поводу дупликации генов проистекает из того факта, что с таким количеством секвенированных геномов, которые сейчас доступны, исследователи имеют более точные оценки того, как часто гены дублируются, и эти показатели чрезвычайно высоки. Например, более 100 генов дублируются в геноме человека за 1 миллион лет (Hahn et al ., 2007a). Это означает, что процентная доля генома, на которую влияют различия в количестве генов (по оценкам, 6%), вносит больший вклад в различия между людьми и шимпанзе, чем однонуклеотидные различия между ортологическими последовательностями (по оценкам, 1,5% ). Высокие показатели (17 генов на 1 миллион лет) также были оценены у мух (Hahn et al.., 2007б). Дополнительный интерес вызывает осознание того, что дупликации происходят так часто, что особи одного и того же вида сильно различаются по содержанию ДНК и количеству генов (т. Е. Многие дупликации являются полиморфными и вносят вклад в индивидуальные различия ). Подсчитано, что в среднем два человека будут различаться примерно на 5 мегабаз информации.
Неожиданно в геномах было описано несколько тенденций к дупликации в отношении эволюции половых хромосом . Многие новые мужские гены происходят из Y-хромосом видов. Некоторые из этих мужских генов организованы в семьи, которые претерпевают генную конверсию, чтобы избежать дегенерации Y-хромосомы. Гены мужской зародышевой линии также могут дублироваться из Х-хромосомы посредством ретропозиции (Betrán et al ., 2002; Emerson et al ., 2004; Lahn et al ., 2001; Rozen et al ., 2003). Эти данные показывают, что расположение и организация генома имеют значение для происхождения и функции генов.
Хромосомные дупликации в эволюции генов
Когда ген дублируется в том же месте или даже в другом, он создает локус с последовательностью и значением. То есть значимая последовательность. Если так и будет, то это будет дубликат своего родительского гена.
Но он может не подвергаться такому же избирательному давлению, как родительский ген, и может мутировать. Сумма этих изменений иногда может привести к появлению новой функции. Уже ген тоже будет новым геном.
Дупликация предкового локуса глобина, например, привела в эволюции к появлению семейства глобинов. Последующие транслокации и последовательные дупликации привели к тому, что семья пополнилась новыми членами, выполняющими ту же функцию, но подходящими для других условий.
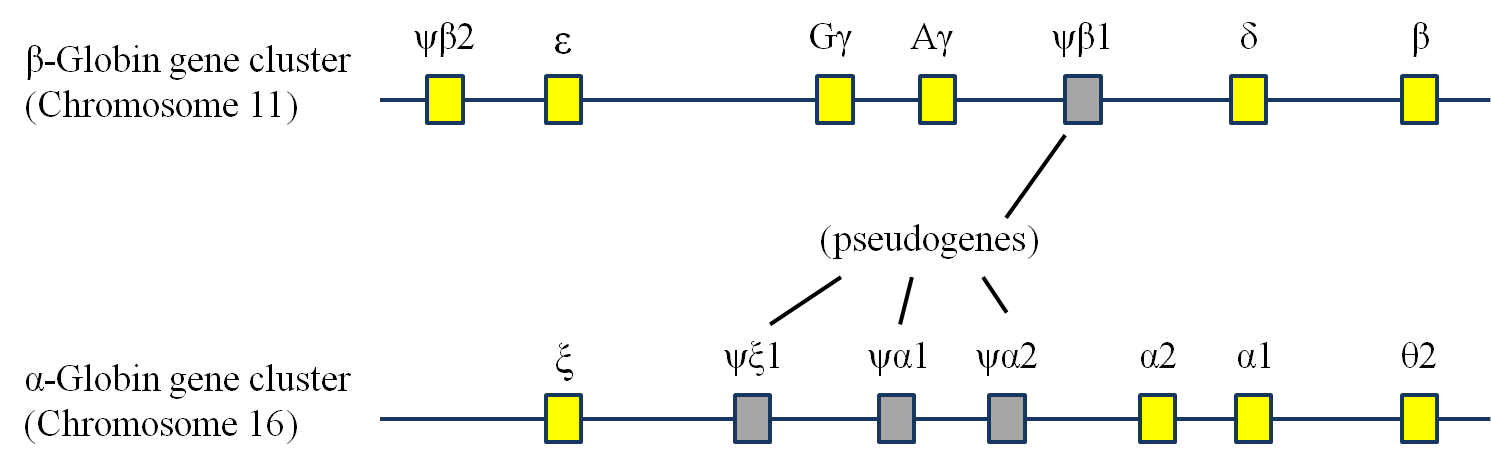
Семейство генов глобинов. Yuhrt, через Wikimedia Commons.
Как эволюционное событие
Эволюционная судьба повторяющихся генов
Скорость дупликации гена
Сравнение геномов показывает, что дупликации генов обычны у большинства исследованных видов. На это указывает переменное количество копий (вариация количества копий) в геноме человека или плодовой мушки. Однако было трудно измерить скорость, с которой происходит такое дублирование. Недавние исследования дали первую прямую оценку скорости дупликации генов по всему геному у C. elegans , первого многоклеточного эукариота, для которого такие оценки стали доступны. Уровень дупликации генов у C. elegans составляет порядка 10-7 дупликаций на ген / поколение, то есть в популяции из 10 миллионов червей у одного будет дупликация гена на поколение. Эта скорость на два порядка больше, чем скорость спонтанной точечной мутации на нуклеотидный сайт у этого вида. В более ранних (непрямых) исследованиях сообщалось о степени локус-специфической дупликации у бактерий, дрозофилы и людей в диапазоне от 10 -3 до 10 -7 / ген / поколение.
Неофункционализация
Дупликации генов — важный источник генетической новизны, которая может привести к эволюционным инновациям. Дупликация создает генетическую избыточность, при которой вторая копия гена часто свободна от селективного давления, то есть ее мутации не оказывают вредного воздействия на организм-хозяин. Если одна копия гена подвергается мутации, которая влияет на ее исходную функцию, вторая копия может служить «запасной частью» и продолжать правильно функционировать. Таким образом, гены-дубликаты накапливают мутации быстрее, чем функциональные гены-единственные копии, на протяжении поколений организмов, и у одной из двух копий может развиться новая и отличная функция. Некоторыми примерами такой неофункционализации являются очевидная мутация дублированного пищеварительного гена в семействе ледяной рыбы в ген антифриза и дупликация, приводящая к новому гену змеиного яда и синтезу 1-бета-гидрокситестостерона у свиней.
Считается, что дупликация генов играет важную роль в эволюции ; такую позицию придерживаются представители научного сообщества более 100 лет. Сусуму Оно был одним из самых известных разработчиков этой теории в своей классической книге « Эволюция путем дупликации генов» (1970). Оно утверждал, что дупликация генов — самая важная эволюционная сила с момента появления универсального общего предка . Основные случаи дупликации генома могут быть довольно частыми. Считается, что весь геном дрожжей подвергся дупликации около 100 миллионов лет назад. Растения — самые плодовитые дупликаторы генома. Например, пшеница — гексаплоид (разновидность полиплоида ), что означает, что у нее шесть копий своего генома.
Субфункционализация
Другая возможная судьба повторяющихся генов заключается в том, что обе копии в равной степени могут накапливать дегенеративные мутации, если любые дефекты дополняются другой копией. Это приводит к нейтральной модели «субфункционализации» или модели DDC (дупликация-дегенерация-комплементация), в которой функциональность исходного гена распределяется между двумя копиями. Ни один из генов не может быть потерян, поскольку оба теперь выполняют важные неизбыточные функции, но в конечном итоге ни один из них не может достичь новых функций.
Субфункционализация может происходить через нейтральные процессы, в которых мутации накапливаются без вредных или положительных эффектов. Однако в некоторых случаях может происходить субфункционализация с явными адаптивными преимуществами. Если предковый ген является плейотропным и выполняет две функции, часто ни одна из этих двух функций не может быть изменена, не затрагивая другую функцию. Таким образом, разделение наследственных функций на два отдельных гена может позволить адаптивную специализацию подфункций, тем самым обеспечивая адаптивное преимущество.
Потеря
Часто в результате геномные вариации гена приводит к дозированному зависимым неврологическим расстройствам , таким как Ретта как синдром и болезнь Пелицеуса-Мерцбахер . Такие вредные мутации, вероятно, будут утеряны в популяции и не будут сохранены или не будут развиваться в новых функциях. Однако многие дупликации на самом деле не являются вредными или полезными, и эти нейтральные последовательности могут быть потеряны или могут распространяться по популяции посредством случайных колебаний через генетический дрейф .
Хромосомные дупликации в эволюции видов
В организме дублирование гена приводит к генерации копии, называемой геном paralog. Хорошо изученным является случай генов глобина, упомянутых выше. Одним из самых известных глобинов является гемоглобин.
Очень трудно представить, что удвоится только кодирующая область гена. Следовательно, каждый ген paralog связан с паралогической областью в организме, которая испытывает дублирование.
В ходе эволюции хромосомные дупликации играли важную роль по-разному. С одной стороны, они дублируют информацию, которая может породить новые функции, изменяя гены с предыдущей функцией.




![Дупликация генов. эволюция [классические идеи в свете новых открытий]](https://lesniepolyani.ru/wp-content/uploads/f/1/0/f109e47504594f28e65072c6d36fc22a.png)
С другой стороны, размещение дупликации в другом геномном контексте (например, в другой хромосоме) может генерировать паралог с другой регуляцией. То есть он может генерировать большую адаптационную способность.
Наконец, области обмена путем рекомбинации также создаются, что приводит к большим геномным перестройкам. Это в свою очередь может представлять происхождение событий видообразования в определенных макроэволюционных линиях.
Как возникают гены-сироты
Большинство генов любого биологического вида также встречаются как минимум у одного другого вида. Последовательности у них могут немного отличаться, но они всё же достаточно схожи, чтобы можно было распознать их родство. В результате случайных мутаций цепочки ДНК со временем расходятся по своему строению, но гомологичные гены (произошедшие от одного участка. — Прим. ред.) по-прежнему можно классифицировать по семействам благодаря их сходству. Например, гены молекул гемоглобина у людей и прочих млекопитающих принадлежат к одному семейству.
Сусуму Оно предложил теорию, согласно которой гены с новыми функциями возникают путем дивергенции (от лат. divergium — расхождение, растекание. — Прим. ред.). Он показал, что дупликация генов с последующей мутацией, в результате которой происходит дивергенция двух гомологичных генов, приводит к образованию новых генов.
Сдвиг рамки считывания
К генным относятся мутации обусловленные сдвигом рамки считывания, когда происходит изменение количества нуклеотидных пар в составе гена. Это может быть как выпадение, так и вставка одной или нескольких нуклеотидных пар в ДНК. Генных мутаций по типу сдвига рамки считывания больше всего. Наиболее часто они возникают в повторяющихся нуклеотидных последовательностях.
Вставка или выпадение нуклеотидных пар может произойти в следствие воздействия определенных химических веществ, которые деформируют двойную спираль ДНК.
Рентгеновское облучение может приводить к выпадению, т. е. делеции, участка с большим количеством пар нуклеотидов.
Вставки нередки при включении в нуклеотидную последовательность так называемых подвижных генетических элементов, которые могут менять свое положение.
К генным мутациям приводит неравный кроссинговер. Чаще всего он происходит в тех участках хромосом, где локализуются несколько копий одного и того же гена. При этом кроссинговер происходит так, что в одной хромосоме возникает делеция участка. Этот участок переносится на гомологичную хромосому, в которой возникает дупликация участка гена.
Если происходит делеция или вставка числа нуклеотидов не кратного трем, то рамка считывания сдвигается, и трансляция генетического кода зачастую обессмысливается. Кроме того, может возникнуть нонсенс-триплет.
Если количество вставленных или выпавших нуклеотидов кратно трем, то, можно сказать, сдвиг рамки считывания не происходит. Однако при трансляции таких генов в пептидную цепь будут включены лишние или утрачены значащие аминокислоты.
Механизмы дублирования
Внематочная рекомбинация
Дупликации возникают в результате события, называемого неравным кроссинговером, которое происходит во время мейоза между смещенными гомологичными хромосомами. Вероятность того, что это произойдет, зависит от степени распределения повторяющихся элементов между двумя хромосомами. Продуктами этой рекомбинации являются дупликация в месте обмена и реципрокная делеция. Эктопическая рекомбинация обычно опосредуется сходством последовательностей в точках дублирования, которые образуют прямые повторы. Повторяющиеся генетические элементы, такие как мобильные элементы, предлагают один источник повторяющейся ДНК, которая может облегчить рекомбинацию, и они часто обнаруживаются в точках разрыва дупликации у растений и млекопитающих.
Схема участка хромосомы до и после события дупликации
Проскальзывание репликации
Проскальзывание репликации — это ошибка репликации ДНК, которая может приводить к дублированию коротких генетических последовательностей. Во время репликации ДНК-полимераза начинает копировать ДНК. В какой-то момент в процессе репликации полимераза отделяется от ДНК, и репликация останавливается. Когда полимераза повторно присоединяется к цепи ДНК, она выравнивает реплицирующую цепь в неправильном положении и случайно копирует один и тот же участок более одного раза. Проскальзыванию репликации также часто способствуют повторяющиеся последовательности, но для этого требуется лишь несколько оснований сходства.
Ретротранспозиция
Ретротранспозоны , в основном L1 , иногда могут действовать на клеточную мРНК. Транскрипты обратно транскрибируются в ДНК и вставляются в случайное место в геноме, создавая ретрогены. Результирующая последовательность обычно не имеет интронов и часто содержит поли-последовательности, которые также интегрированы в геном. Многие ретрогены обнаруживают изменения в регуляции генов по сравнению с последовательностями их родительских генов, что иногда приводит к новым функциям.
Анеуплоидия
Анеуплоидия возникает, когда нерасхождение одной хромосомы приводит к аномальному количеству хромосом. Анеуплоидия часто опасна и у млекопитающих регулярно приводит к самопроизвольным абортам (выкидышам). Некоторые анеуплоидные особи жизнеспособны, например трисомия 21 у человека, которая приводит к синдрому Дауна . Анеуплоидия часто изменяет дозировку генов вредным для организма образом; поэтому маловероятно, что он будет распространяться среди населения.
Полиплоидия
Полиплоидия или дупликация всего генома — это продукт нерасхождения во время мейоза, что приводит к появлению дополнительных копий всего генома. Полиплоидия является обычным явлением у растений, но также встречается и у животных, с двумя раундами дупликации всего генома ( событие 2R ) в линии позвоночных, ведущих к человеку. Это также произошло в дрожжах гемиаскомицетов ~ 100 млн лет назад.
После дупликации всего генома наступает относительно короткий период нестабильности генома, обширная потеря генов, повышенные уровни нуклеотидных замен и перестройка регуляторной сети. Кроме того, важную роль играют эффекты дозировки генов. Таким образом, большинство дубликатов теряется в течение короткого периода времени, однако значительная часть дубликатов выживает. Интересно, что гены, участвующие в регуляции, преимущественно сохраняются. Более того, сохранение регуляторных генов, в первую очередь Hox-генов , привело к адаптивным инновациям.
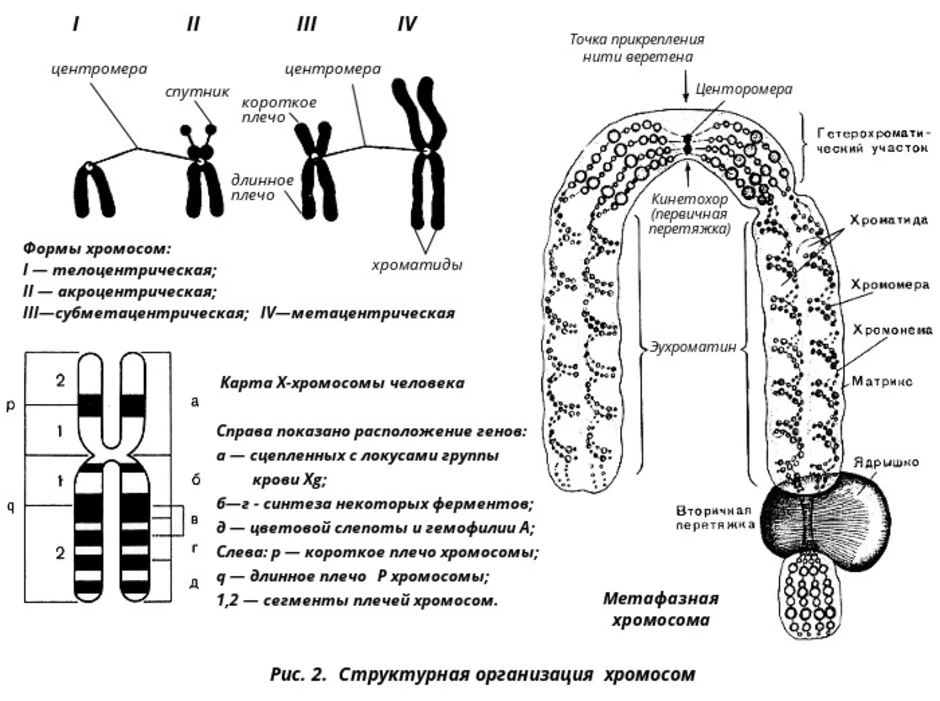


Быстрая эволюция и функциональная дивергенция наблюдаются на уровне транскрипции дублированных генов, обычно за счет точечных мутаций в коротких мотивах связывания факторов транскрипции. Более того, быстрая эволюция мотивов фосфорилирования белков, обычно встроенных в быстро эволюционирующие внутренне неупорядоченные области, является еще одним фактором, способствующим выживанию и быстрой адаптации / неофункционализации повторяющихся генов. Таким образом, кажется, существует связь между регуляцией генов (по крайней мере, на посттрансляционном уровне) и эволюцией генома.
Полиплоидия также является хорошо известным источником видообразования, поскольку потомство, которое имеет другое количество хромосом по сравнению с родительскими видами, часто неспособно скрещиваться с неполиплоидными организмами. Считается, что дупликации всего генома менее вредны, чем анеуплоидия, поскольку относительные дозы отдельных генов должны быть одинаковыми.
Тандемное дублирование экзонов
Тандемное дублирование экзонов соответствует внутреннему дублированию экзона в одном и том же гене. Этот тип дупликации является важным источником инноваций, поскольку он позволяет генерировать новые fs в одном и том же белке. Всесторонний анализ геномов Homo sapiens , Drosophila melanogaster и червя Caenorhabditis elegans показал 12291 случай тандемной дупликации экзонов. Анализ интронных областей также выявил 4660 не охарактеризованных дублированных экзонов. Среди этих потенциальных экзонов 35,1% обнаружены в базах данных EST, что подтверждает их потенциальную роль.
Что происходит с новыми генами?
Все эти новые последовательности увеличивают сложность и разнообразие геномов. Как и в случае любой мутации , когда новые гены закрепляются в геноме, они увеличивают различия между видами и служат сырьем для эволюции (Ohno, 1970). Это легко увидеть в случае дупликации генов. Дублирование гена приводит к образованию двух или более копий гена: одной, которая может поддерживать свою первоначальную функцию в организме , а другой (-ых) можно «поиграть», чтобы взять на себя новые функции. Как следствие, новые дубликаты являются основным источником нововведений в геноме и часто развиваются при положительном отборе , при котором происходят быстрые изменения в белке, кодируемом новым геном, чтобы получить новую функцию (Presgraves, 2005). Этот процесс называетсянеофункционализация нового гена.
Другие возможные исходы после дупликации включают потерю гена или псевдогенизацию; поддержание обоих генов как способ увеличения экспрессии или поддержания множественных вариантов у индивидуумов (по существу «фиксация» гетерозиготности ); или возникновение субфункционализации (то есть возникновение взаимно дополняющих нейтральных выводящих мутаций, так что оба гена необходимо сохранить в геноме ). Субфункционализация — интересное явление, потому что оно начинается с разделения функций, но может заложить основу для специализации (Torgerson & Singh, 2004). Также возможны некоторые смешанные исходы (например, субфункционализация с последующей неофункционализацией и субнеофункционализацией) (He & Zhang, 2005).
Одним из непредвиденных последствий дупликации и потери генов является то, что эти события могут стать причиной несовместимости между видами. Было показано, что дупликации и потери играют роль в распаде гибридов и снижении приспособленности потомков к спариванию между генетически дифференцированными популяциями. Таким образом, эти процессы могут вносить вклад в процесс видообразования (Masly et al ., 2006).