

Содержание
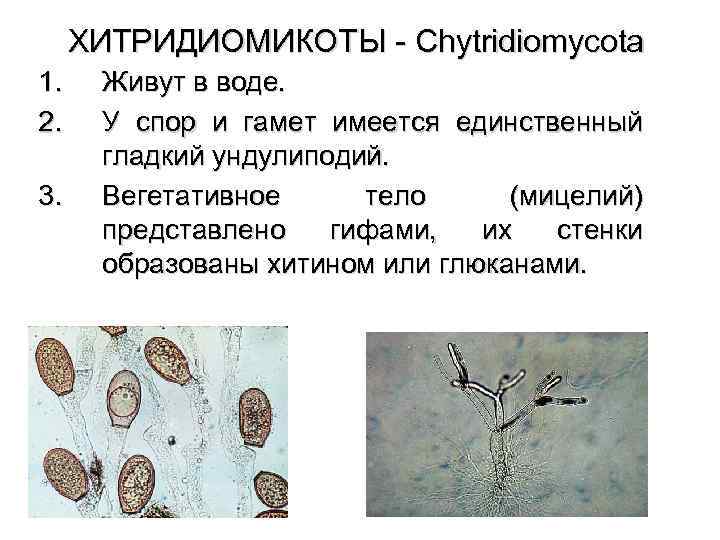
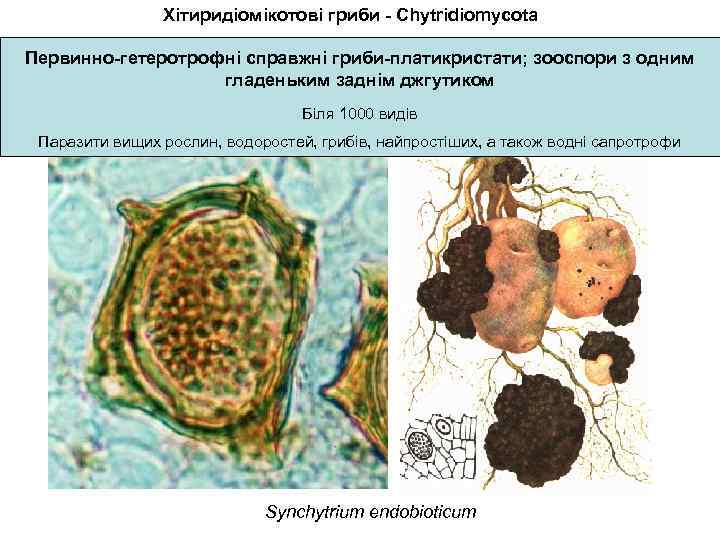
Хитридиомикот это тип, который группирует микроскопические грибы со спорами жгутиков. Возможно, они являются предковой группой высших грибов. Его систематика сложна из-за пластичности форм в разных условиях выращивания.
Они питаются абсорбцией. Существуют сапрофиты и паразиты, некоторые из которых являются патогенами для сельскохозяйственных культур, таких как картофель, а другие наносят ущерб популяциям амфибий. Они обитают в почве, а также в пресных и соленых водах. Некоторые облигатно-анаэробные виды населяют пищеварительный тракт травоядных.
Они размножаются бесполым путем с помощью зооспор, вызывая гаплоидную фазу, в которой образуются реснитчатые гаметы. Затем с помощью плазмогамии и кариогамии образуется диплоидная фаза, которая дает начало споротой.
Среды обитания
Читриды водные грибы, хотя те, что процветают в капиллярной сети вокруг частиц почвы, обычно считаются наземными. Зооспора — это в первую очередь средство тщательного исследования небольшого объема воды в поисках подходящего субстрата, а не средство распространения на большие расстояния.
Хитриды были изолированы из различных водных сред обитания, включая торф, болота, реки, пруды, родники и канавы, а также из наземных сред обитания, таких как кислые почвы, щелочные почвы, почвы умеренных лесов, почвы тропических лесов, почвы Арктики и Антарктики. Это привело к убеждению, что многие виды хитридов распространены повсеместно и космополитичны. Однако недавняя таксономическая работа продемонстрировала, что этот вездесущий и космополитичный морфовид скрывает загадочное разнообразие на генетическом и ультраструктурном уровнях. Сначала считалось, что водные хитриды (и другие зооспорические грибы) в основном активны осенью, зимой и весной. Однако недавняя молекулярная инвентаризация озер летом показывает, что хитриды являются активной и разнообразной частью микробного сообщества эукариот.
Одной из наименее ожидаемых наземных сред, в которых процветает хитрид, являются перигляциальные почвы. Популяция видов Chytridiomycota может поддерживаться, даже если в этих замерзших регионах отсутствует растительная жизнь из-за большого количества воды в перигляциальной почве и пыльцы, поднимающейся из-под границы леса.
Классы хитридиомицеты, оомицеты и зигомицеты
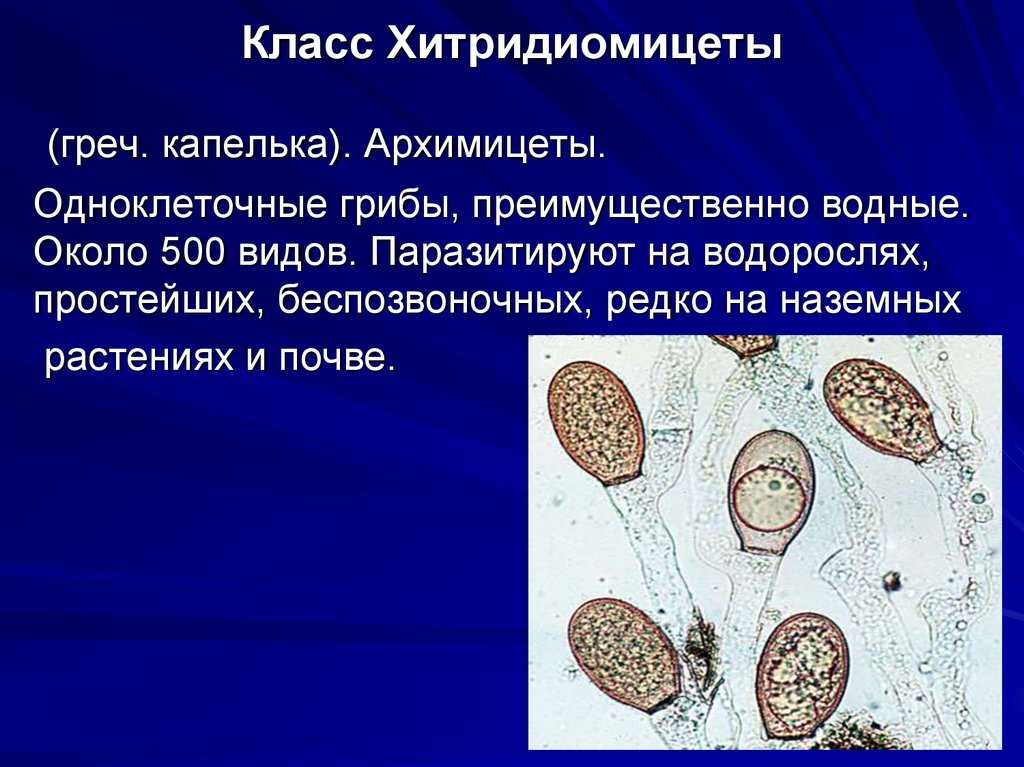
Положение этого отдела в системе грибов спорно, но данные геносистематики подтверждают, что это наиболее древняя группа грибов, единственная, имеющая в жизненных циклах подвижные стадии, примитивно устроенный таллом.
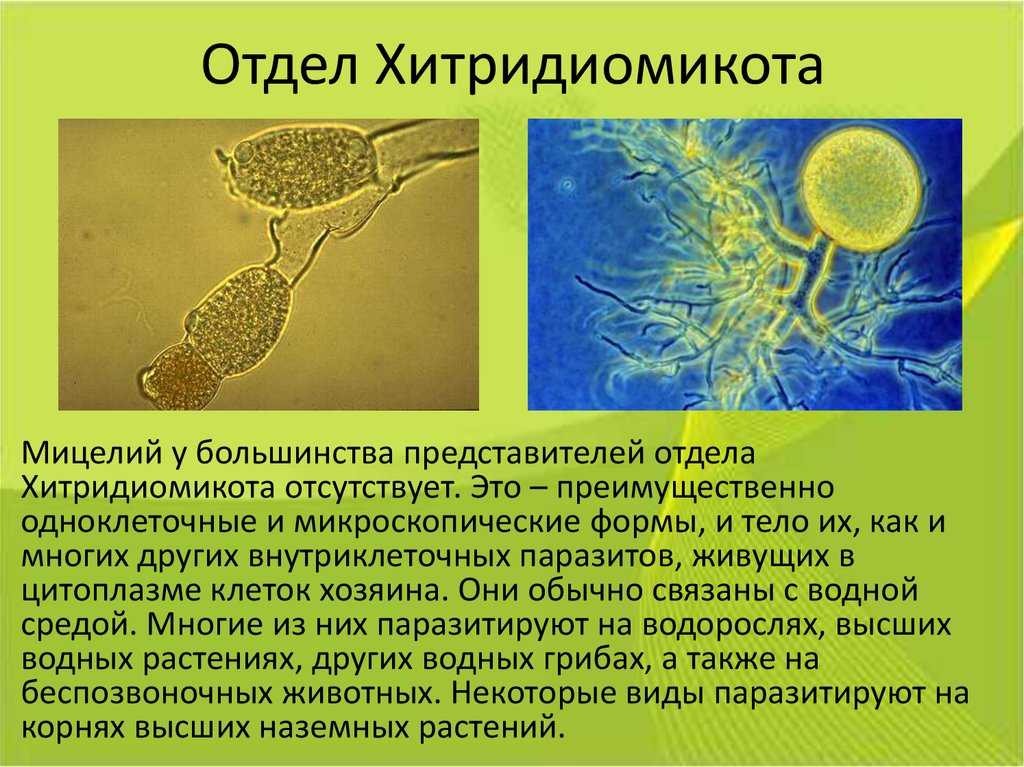
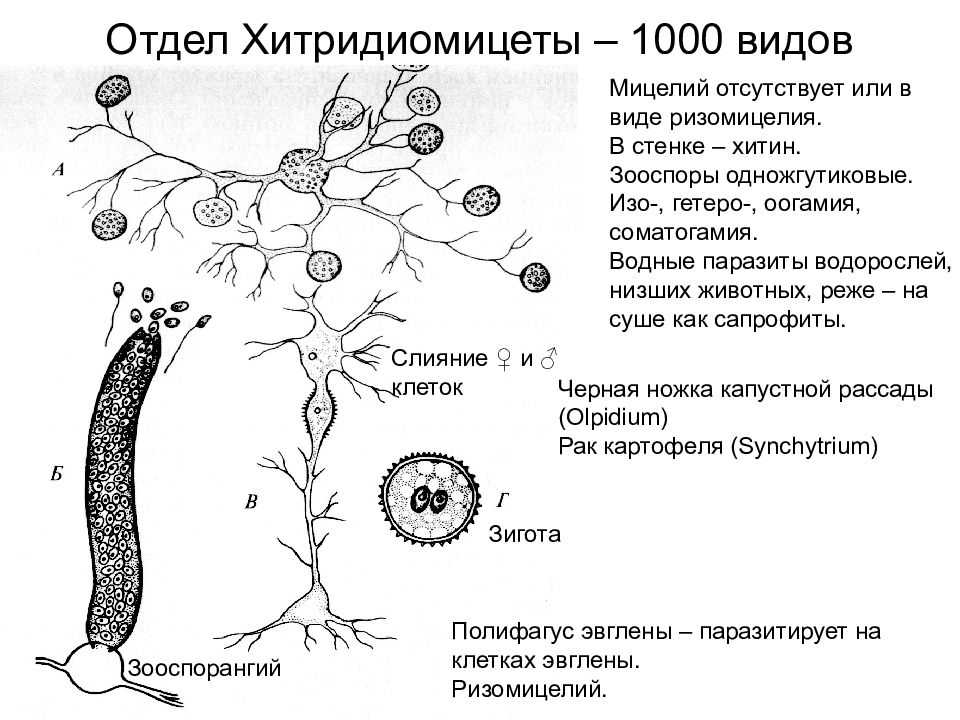
Вегетативное тело у большинства видов развито слабо.
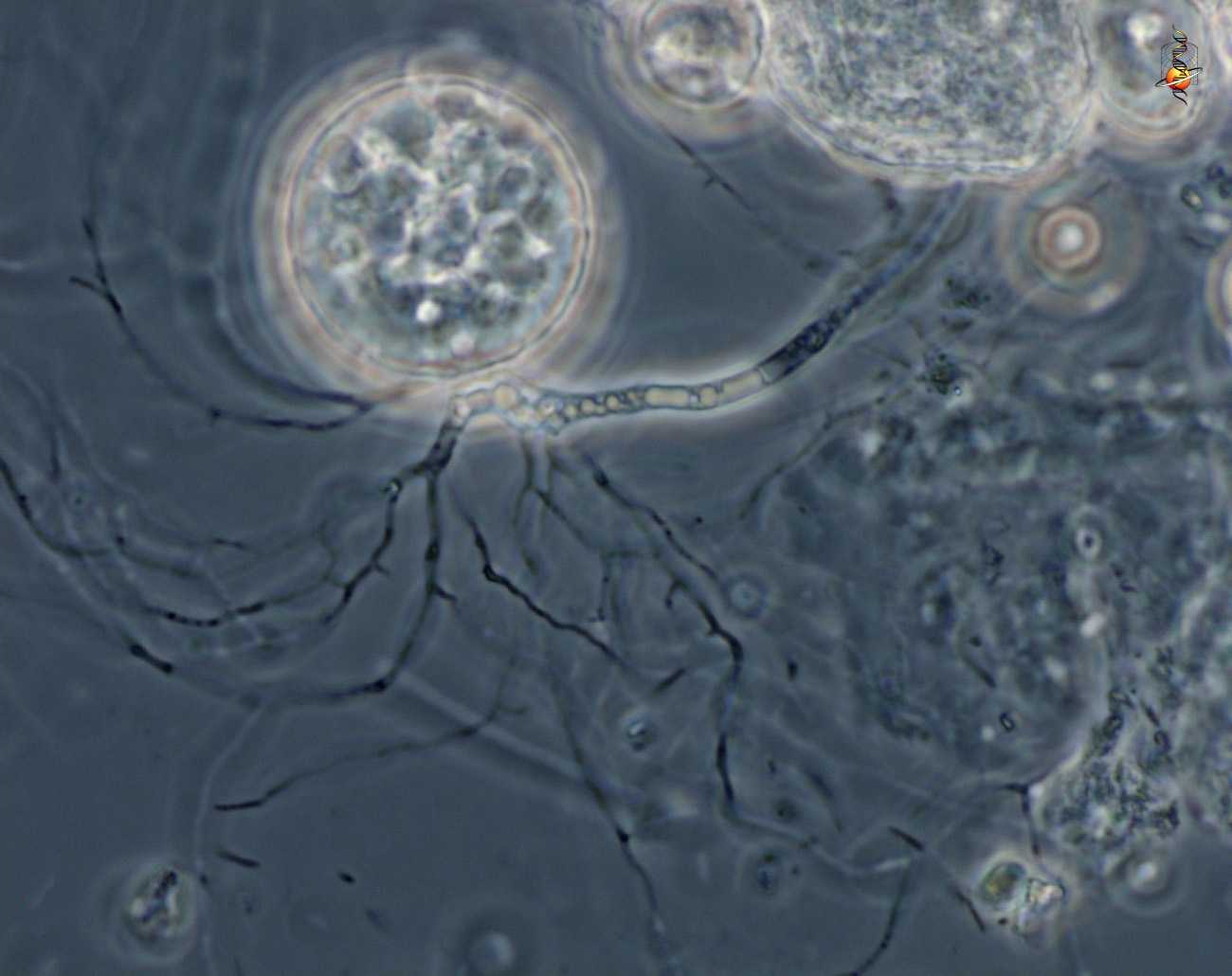
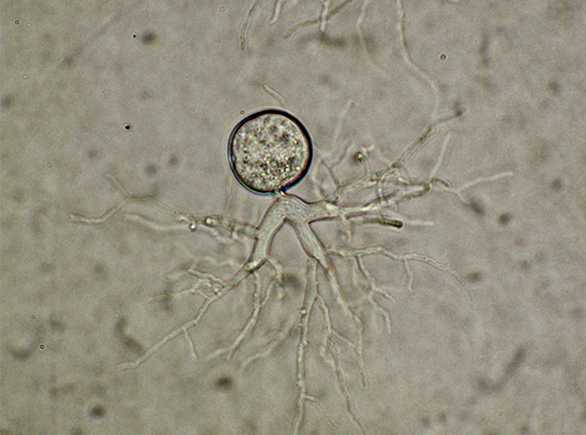
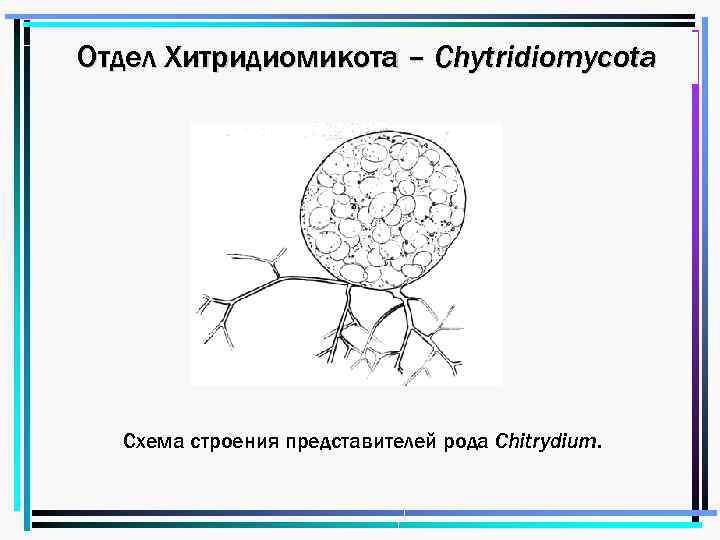
Чаще таллом одноклеточный, может быть с ризомицелием. У более высокоорганизованных (порядки моноблефаридальные — Monoblepharidales, бластокладиальные — Blastocladiales) вегетативное тело в виде многоядерного разветвленного неклеточного мицелия. Клеточная оболочка хитиново-глюкановая.
У большинства видов талломы с одним ядром в центральной части и без ядер в ризомицелии, но у некоторых представителей талломы многоядерные со специальными структурами — собирательными клетками.
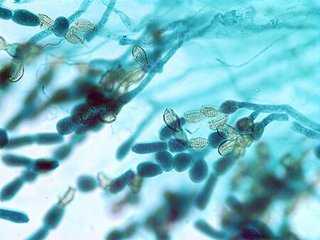
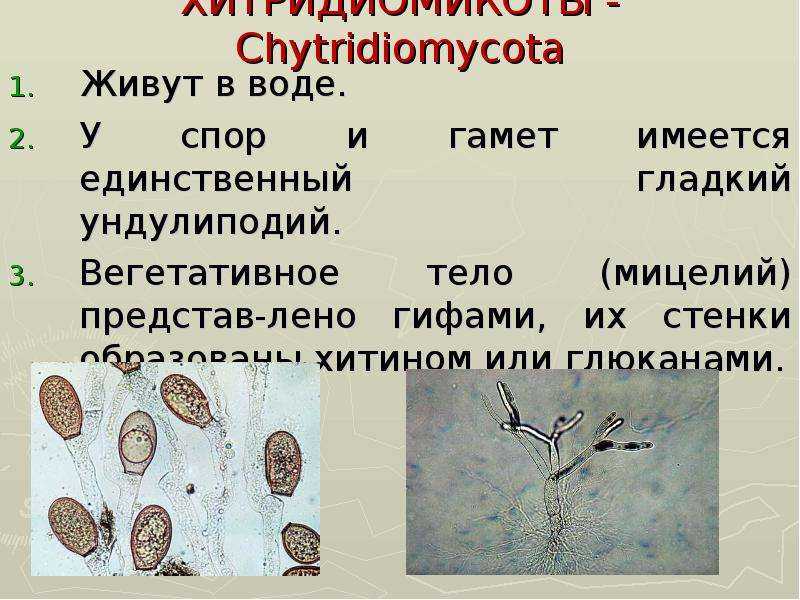
Бесполое размножение осуществляется с помощью зооспор, имеющих один гладкий бичевидный жгутик, прикрепленный к заднему концу споры.
Оседая на субстрат, зооспоры теряют жгутики и разными способами превращаются в вегетативное тело гриба:
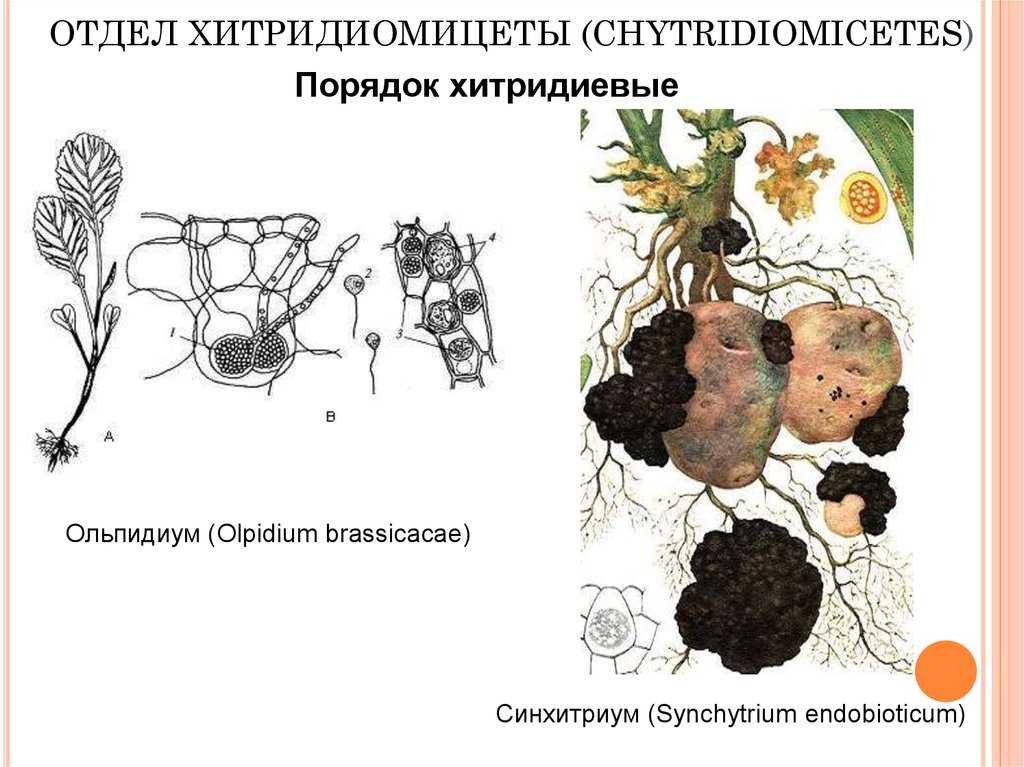
- — у эндопаразитов содержимое зооспор переходит в клетку растения по специальному каналу;
- — у эктопаразитов зооспоры покрываются оболочкой и остаются сверху, а внутрь клетки проникает ризомицелий;
- — у сапротрофов тело образуется внутри и снаружи субстрата.
Зооспоры образуются в шаровидных или грушевидных зооспорангиях, из которых выходят через пору на конце выводковой трубки или через отверстие, открывающееся специальной крышечкой.
У более примитивных организмов одноклеточное вегетативное тело превращается в спорангий, у более высокоорганизованных видов спорангии образуются из части вегетативного тела.
Характерно многообразие типов полового процесса (хотя у многих представителей половой процесс не является достоверным).
Это может быть изогамия, гетерогамия, оогамия и даже соматогамия. Продукт полового размножения — покоящиеся зиготы, покрытые оболочкой.
У ряда хитридиомицетов прослеживается два типа чередования бесполого и полового поколений (как у водорослей): изоморфная и гетероморфная смена генераций.
В циклах развития преобладает гаплоидная стадия. У большинства видов — мейоз зиготический.
Обитают в воде или во влажной наземной среде. Паразитируют на водорослях, цветковых растениях, низших животных и грибах. Некоторые виды могут быть сапротрофами.
Классификация.
Отдел включает один класс Chytridiomycetes (хитридиомицеты) (4…6-го порядков). Выделение порядков в настоящее время основано на ультраструктуре зооспор
Обращается внимание на такие признаки, как строение жгутикового аппарата, форма и расположение ядра, митохондрий, рибосом, липидных глобул и др
Жизненный цикл и план тела
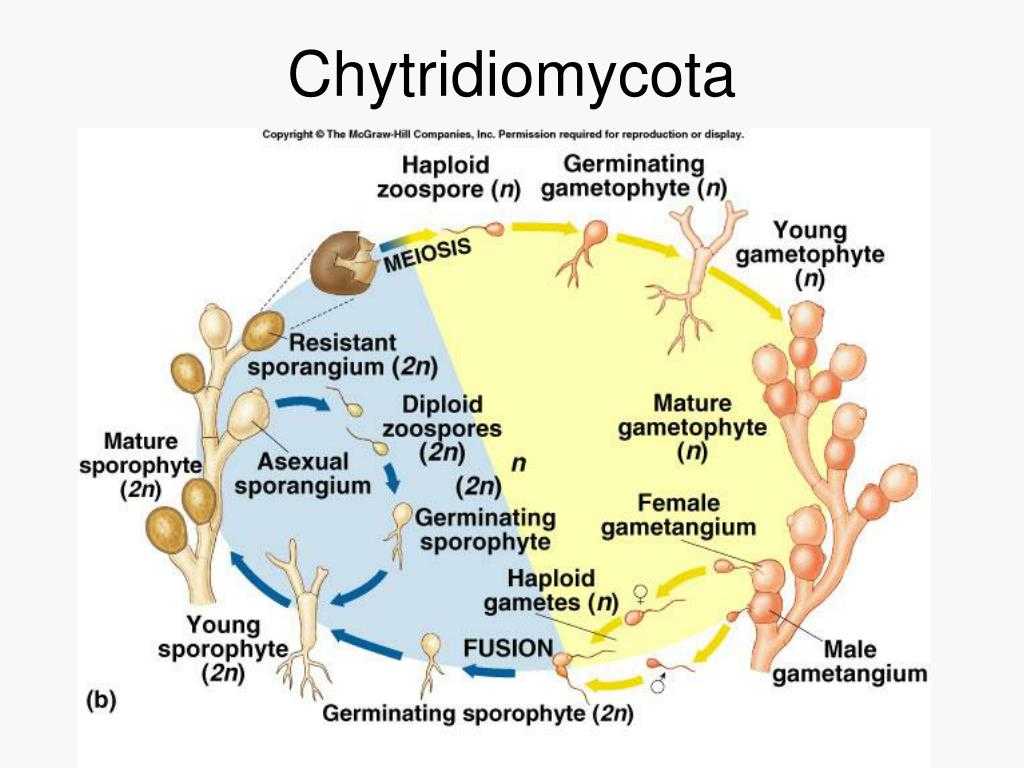
Chytridiomycota необычны среди грибов тем, что они размножаются зооспорами. Для большинства представителей Chytridiomycetes половое размножение неизвестно. Бесполое размножение происходит за счет высвобождения зооспор (предположительно), полученных через митоз.

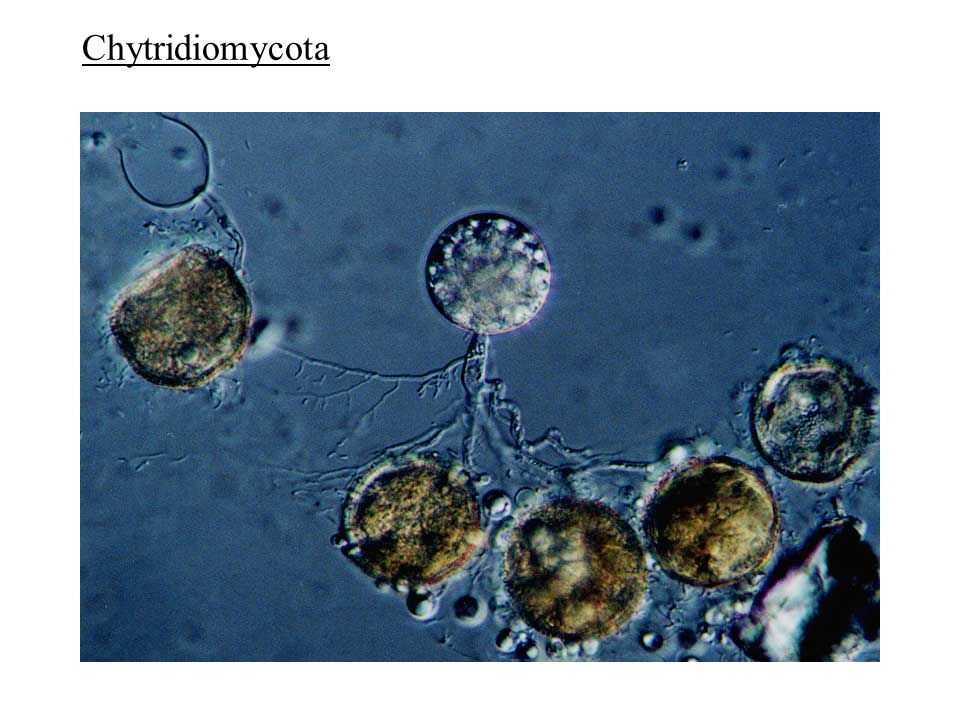
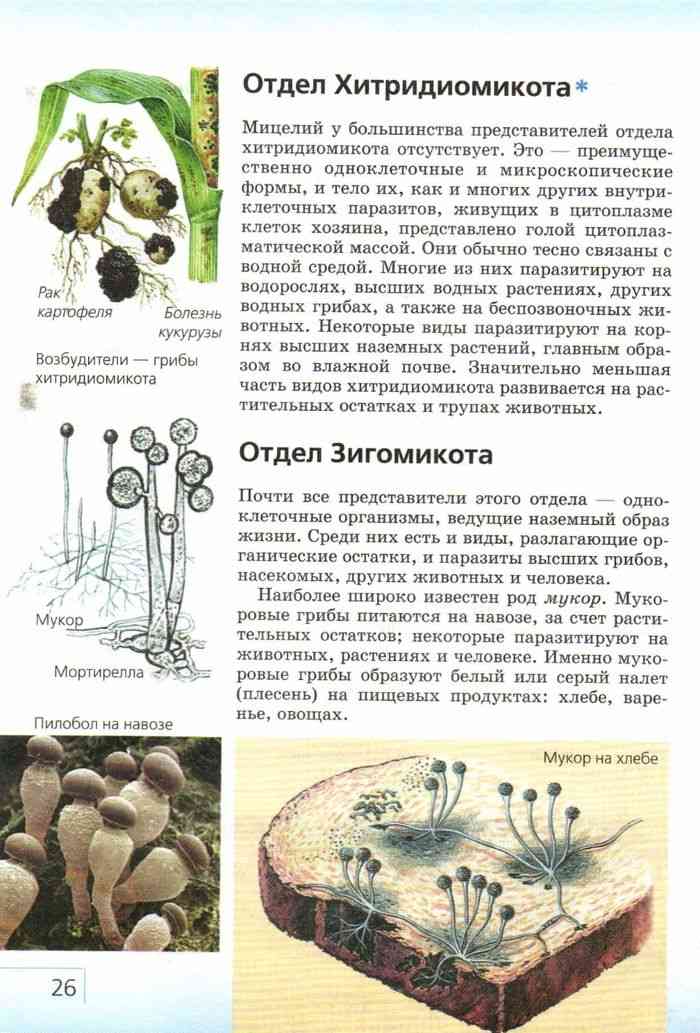
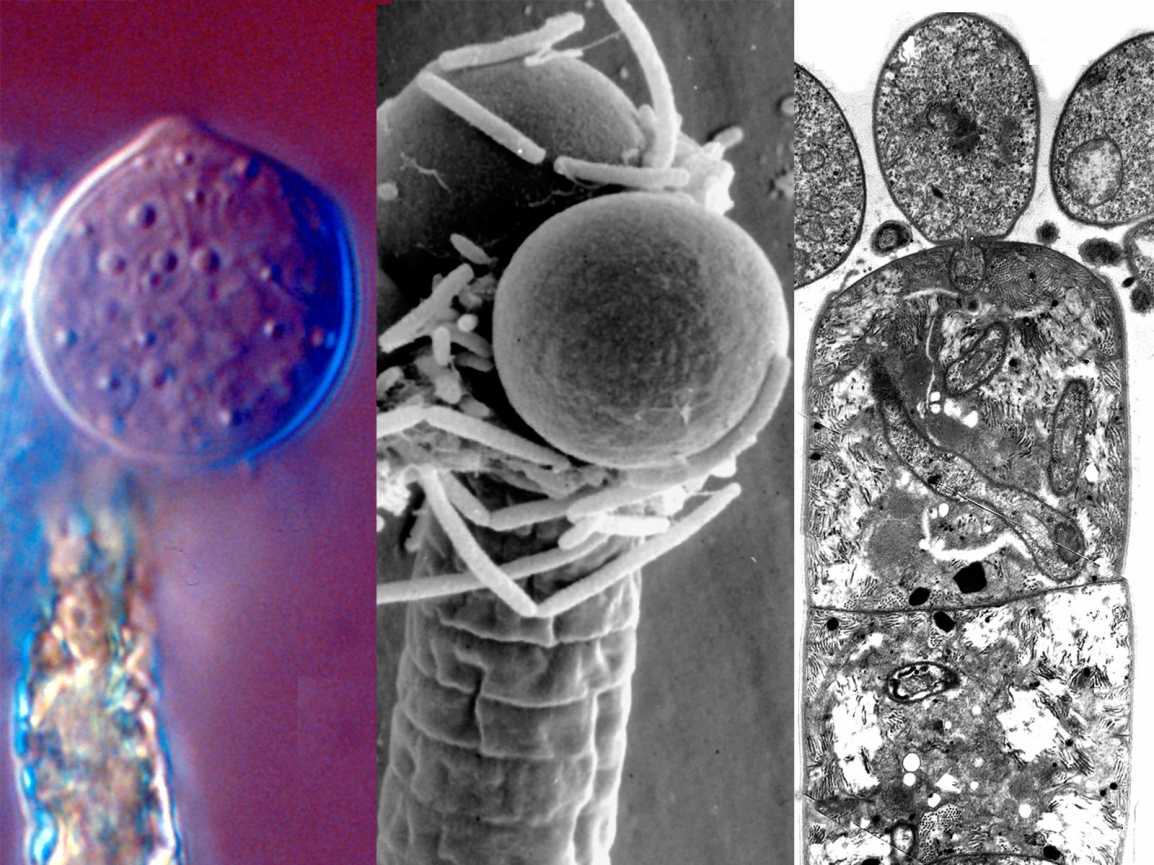
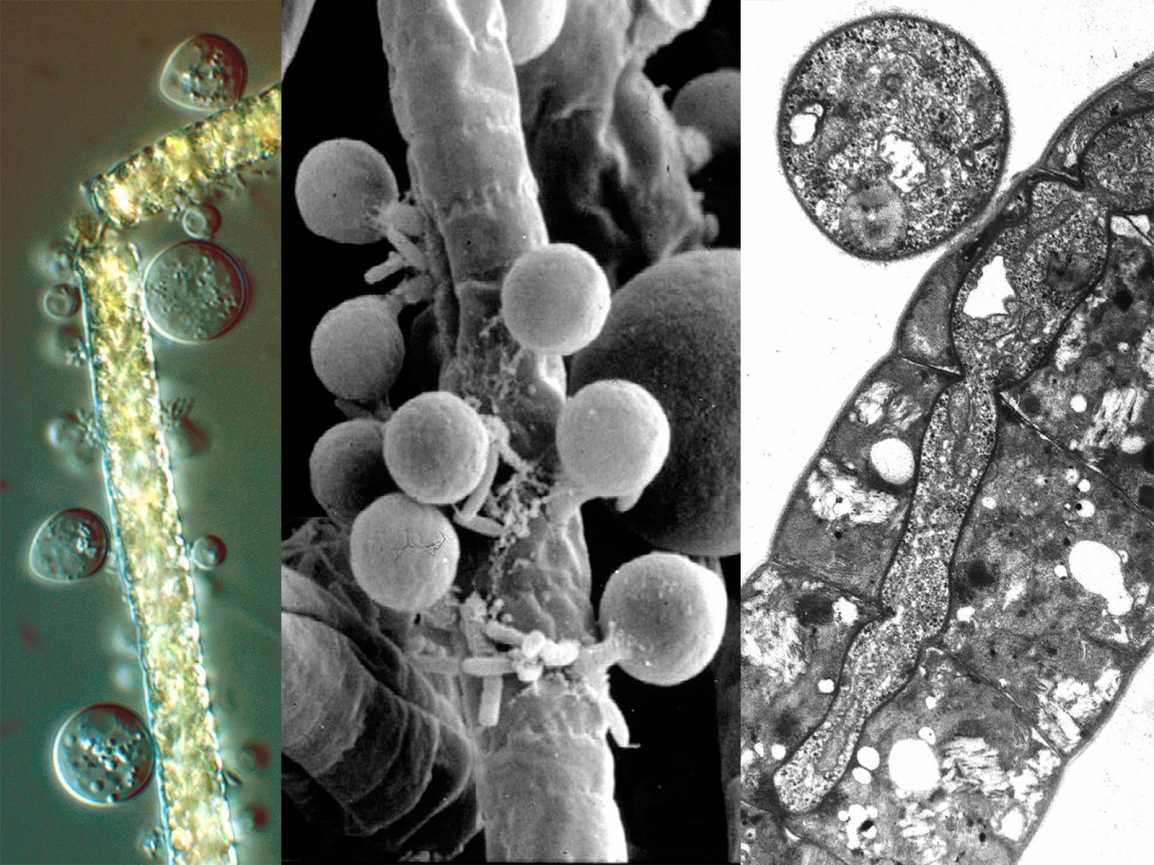

Там, где это было описано, половое размножение Chytridomycetes происходит различными способами. Принято считать, что образующаяся зигота образует покоящуюся спору, которая действует как средство выживания в неблагоприятных условиях. У некоторых членов половое размножение достигается за счет слияния изогамет (гаметы одинакового размера и формы). В эту группу входят известные патогены растений. Синхитрий. Некоторые водорослевые паразиты практикуют оогамия: подвижная мужская гамета прикрепляется к неподвижной структуре, содержащей женскую гамету. В другой группе двое слоевища производят трубки, которые сливаются и позволяют гаметам встречаться и сливаться. В последней группе встречаются и сливаются ризоиды совместимых штаммов. Оба ядра мигрируют из зооспорангия в соединенные ризоиды, где они сливаются. Образовавшаяся зигота прорастает в покоящуюся спору.
Половое размножение распространено и хорошо известно среди представителей моноблефаридомицетов. Обычно эти читриды практикуют разновидность оогамия: самец подвижен, а самка неподвижна. Это первое проявление оогамии в королевстве Грибки. Вкратце, моноблефы образуют оогонии, дающие начало яйцам, и антеридии, дающие начало мужским гаметам. После оплодотворения зигота становится инцистированной или подвижной ооспорой, которая в конечном итоге становится покоящейся спорой, которая позже прорастет и даст начало новым зооспорангиям.
После освобождения от проросших покоящихся спор зооспоры ищут подходящий субстрат для роста, используя хемотаксис или фототаксис. Некоторые виды инцизируют и прорастают непосредственно на субстрате; другие инцизируют и прорастают на небольшом расстоянии. После прорастания ферменты, высвобождаемые из зооспор, начинают расщеплять субстрат и использовать его, образуя новый слоевище. Талли ценоцитарный и обычно не соответствуют действительности мицелий (имея ризоиды вместо).
Хитриды имеют несколько различных моделей роста. Некоторые из них голокарпические, что означает, что они производят только зооспорангий и зооспоры. Другие являются евкарпическими, что означает, что они создают другие структуры, такие как ризоиды, помимо зооспорангия и зооспор. Некоторые хитриды моноцентрические, то есть одна зооспора дает начало одному зооспорангии. Другие полицентричны, то есть одна зооспора дает начало множеству зооспорангиев, связанных ризомицелием. Ризоиды не имеют ядер, в отличие от ризомицелия.
Рост продолжается до тех пор, пока новая партия зооспор не будет готова к выпуску. Хитриды обладают разнообразным набором механизмов высвобождения, которые могут быть сгруппированы в широкие категории «неперкулирующие» и «неперкулирующие». Жаберные выделения включают полное или неполное отслоение крышки, похожей на крышку, называемую крышечкой, которая позволяет зооспорам выйти из спорангия. Недостаточные хитриды выпускают свои зооспоры через поры, щели или сосочки.
Экологические функции
Хитридовые паразиты морских диатомовых водорослей. (А) Хитридовые спорангии на Pleurosigma sp. Белая стрелка указывает на поры сверхчувствительного отделяемого. (B) Ризоиды (белая стрелка) распространяются на хозяина диатомовой водоросли. (C) Агрегаты хлорофилла, локализованные в местах инфекции (белые стрелки). (D и E) Одиночные хозяева, несущие несколько зооспорангиев на разных стадиях развития. Белая стрелка на панели E выделяет ветвящиеся ризоиды. (F) Эндобиотические хитридоподобные спорангии внутри створки диатомовых водорослей. Штанги = 10 мкм.
Пеннатная диатомовая водоросль из арктического плавильного пруда , инфицированная двумя хитридоподобными грибковыми патогенами спорангиев (окрашена в красный цвет).
Batrachochytrium dendrobatidis
Хитрид Batrachochytrium dendrobatidis ответственен за хитридиомикоз , болезнь земноводных. Обнаруженная в 1998 году в Австралии и Панаме, эта болезнь, как известно, убивает большое количество земноводных , и была предложена в качестве основной причины сокращения численности земноводных во всем мире . Было обнаружено, что вспышки грибка ответственны за гибель большей части популяции опрысканной жабы Киханси в ее естественной среде обитания в Танзании , а также за исчезновение золотой жабы в 1989 году. Хитридиомикоз также был причастен к предполагаемому исчезновению южной желудочной лягушки-матки. , последний раз видели в дикой природе в 1981 году, и северную желудочную лягушку-выводок, последний раз отмеченную в дикой природе в марте 1985 года. во время его тиражирования.
Недавние исследования показали, что небольшое повышение уровня соли может вылечить хитридиомикоз у некоторых видов австралийских лягушек, хотя необходимы дальнейшие эксперименты.
Прочие паразиты
Хитриды в основном поражают водоросли и другие эукариотические и прокариотические микробы. Инфекция может быть настолько серьезной, что ограничивает первичную продукцию в озере. Было высказано предположение, что паразитические хитриды оказывают большое влияние на пищевые сети озер и прудов. Хитриды могут также поражать виды растений; в частности, Synchytrium endobioticum является важным патогеном картофеля .
Сапробы
Возможно, самая важная экологическая функция хитридов — это разложение. Эти вездесущие и космополитические организмы ответственны за разложение тугоплавких материалов, таких как пыльца , целлюлоза , хитин и кератин . Есть также хитриды, которые живут и растут на пыльце, прикрепляя нитевидные структуры, называемые ризоидами, к пыльцевым зернам. В основном это происходит во время бесполого размножения, потому что зооспоры, которые прикрепляются к пыльце, непрерывно воспроизводятся и образуют новые хитриды, которые будут прикрепляться к другим зернам пыльцы для получения питательных веществ. Эта колонизация пыльцы происходит весной, когда в водоемах накапливается пыльца, падающая с деревьев и растений.
Мертвая лягушка с признаками хитридиомикоза ( B. dendrobatidis )
Окаменелости
Раннее окаменелости хитридов из Шотландский Рини черт, а Девонский -возраст lagerstätte с анатомической сохранностью растения и грибки. Среди микрофоссилий встречаются хитриды, сохранившиеся в виде паразиты на риниофиты. Эти окаменелости очень напоминают современный род . Останки голокарпического хитрида были найдены в кремнях из Комбре в центральной Франции, которые относятся к позднему периоду. Визейский. Эти останки были найдены вместе с останками евкарпиков и имеют неоднозначный характер, хотя, как считается, они принадлежат к хитридам. Другие хитридоподобные окаменелости были обнаружены в кремнях из верхних слоев. Пенсильванский в Сент-Этьен Бассейн в Франция, датируемые между 300 и 350 годамима.
Класс Оомицеты (Oomycetes)
(Царство Chromista, отдел Oomycota)
Грибы этого класса характеризуются хорошо развитым, но одноклеточным разветвленным мицелием. Клеточная стенка содержит целлюлозу, хитин отсутствует.
Половой процесс проходит по типу оогамии – женская клетка – оогоний, сливаясь с мужской клеткой – антеридием, превращается в половую покоящуюся клетку – ооспору, которая обычно зимует в тканях растений.
Бесполое размножение у низкоорганизованных видов, ведущих водный образ жизни, осуществляется двухжгутиковыми зооспорами, образующимися в зооспорангиях. Более развитые виды, перешедшие к наземному образу жизни, размножаются с помощьюконидий. Конидии образуются на
Таблица 1 – Распределение грибов и грибоподобных организмов по царствам, основным отделам и классам конидиеносцах, выступающих из пораженных тканей наружу.
Строение конидиеносцев и характер их ветвления является одним из признаков систематического деления оомицетовых грибов.
| Царство Protozoa (Простейшие) | Царство Chromista (Псевдогрибы) | Царство Mycota, Fungi (Настоящие грибы) | |||||||||||
| Отдел Myxomycota (Слизевики) | Отдел Plasmodio-phoroмycota(Плазмодиафоровые) | Отдел Oomycota (Оомицеты) | Отдел Zygomycota (Зигомицеты) | Отдел Chytridio-mycota (Хитридиомицеты) | Отдел Dicaryomycota | Deuteromycetes, Mitosporis fungi, Fungi imperfecti (Несовершенные) | |||||||
| Подотдел Ascomycotina (Сумчатые) | Подотдел Basidiomycotina (Базидиальные) | ||||||||||||
| Класс Myxomycetes | Класс Psasmodiophoromycetes | Класс Oomycetes | Класс Zygomycetes | Класс Chytridiomyces | Класс Archae- ascomy- cetes | Класс Euasco- mycetes | Класс Loculoasco- mycetes | Класс Teliomy- cetes | Класс Ustomy- cetes | Класс Basidio- mycetes | Класс Agono- mycetes | Класс Hypho- mycetes | Класс Coelo- mycetes |
| Размножение: половое — изогамия, бесполое — зооспорами | Спороношения: половое – ооспоры бесполое –зооспоры, конидии | Спороношения: половое –зиго- споры, бесполое –спо- рангиоспоры, конидии | Размножение: половое — изогамия, гетерогамия бесполое — зооспорами | Спороношения: половое — сумкоспоры, бесполое — конидии | Спороношения: половое – базидиоспоры, бесполое — типа конидий | Спороношения: половое — отсутствует, бесполое – конидии или отсутствует | |||||||
| Вегетативное тело — плазмодий | Вегетативное тело – одноклеточный мицелий | Вегетативное тело — плазмодий | Вегетативное тело многоклеточный мицелий | ||||||||||
| Низшие грибы | Высшие грибы | ||||||||||||
По строению мицелия, зооспорангиев, конидий и зооспор класс Оомицеты делится на 5 порядков. В фитопатологии наибольшее значение имеют грибы из порядка пероноспоровых (Peronosporales). Большинство грибов этого порядка являются паразитами высших растений, вызывая заболевания листьев, побегов и в конечном итоге гибель растений.
В порядок пероноспоровые входят три семейства: питиевые (Pythiaceae), пероноспоровые (Peronosporaceae) и альбуговые (Albuginaceae).
Наибольшее значение для лесного и сельского хозяйства имеют грибы из семейства питиевые, включающие роды фитофтора (Phytophthora) и питиум (Pithium). Род фитофтора включает более 70 видов грибов, специализирующихся на разных видах сельскохозяйственных и древесных растений. Гриб Phytophthora omnivora вызывает гниль сеянцев лиственных пород. Особую опасность представляет гриб Ph. infestans, поражающий картофель, томаты и другие пасленовые культуры.
В роде Pithium наибольшее значение для лесного хозяйства имеет гриб P.de baryanum, вызывающий загнивание проростков и полегание сеянцев древесных и кустарниковых пород, он также поражает многие сельскохозяйственные культуры (капусту, свеклу, кукурузу и др.), вызывая почернение оснований стеблей и гибель растений («черная ножка»).
Все представители семейства пероноспоровые (Peronosporaceae) – облигатные паразиты. Зараженные этими грибами растения болеют ложной мучнистой росой или пероноспорозом. Так, грибы из рода Peronospora вызывают ложную мучнистую росу листьев огурцов и других тыквенных. Гриб Plasmopara viticola – является возбудителем одной из наиболее опасных болезней винограда – милдью, а гриб Blapharospora combivora вызывает чернильную болезнь каштана съедобного.
Представители семейства Albuginaceae также являются облигатными паразитами высших растений, вызывая деформацию и вздутия листьев, плодов, стеблей с характерным белым налетом (белая ржавчина). Например, вид Albugo candida поражает растения из семейства крестоцветных.



Классификация
Виды Chytridiomycota традиционно выделяются и классифицируются на основе развития, морфологии, субстрата и метода выделения зооспор. Однако изоляты с одиночными спорами (или изогенные линии) демонстрируют большое количество вариаций по многим из этих характеристик; таким образом, эти признаки не могут использоваться для надежной классификации или идентификации видов. В настоящее время таксономия Chytridiomycota основана на молекулярных данных, ультраструктуре зооспор и некоторых аспектах морфологии и развития слоевища.
В более старом и более узком смысле (здесь не используется) термин «хитриды» относится только к грибам из класса Chytridiomycetes . Здесь термин «хитрид» относится ко всем членам Chytridiomycota.
Хитриды также были включены в число протоктистов , но теперь их регулярно классифицируют как грибы.
В более старых классификациях хитриды, за исключением недавно установленного порядка Spizellomycetales , были помещены в класс Phycomycetes подтипа Myxomycophyta царства Fungi. Ранее они относились к мастигомикотинам как класс Chytridiomycetes. Другие классы Mastigomycotina, Hyphochytriomycetes и Oomycetes , были удалены из грибов и классифицированы как гетероконт псевдогрибки .
Класс Chytridiomycetes насчитывает более 750 видов хитридов, распределенных по десяти отрядам . Дополнительные классы включают Monoblepharidomycetes с двумя отрядами и Hyaloraphidiomycetes с одним отрядом .
Молекулярная филогенетика и другие методы, такие как анализ ультраструктуры , значительно расширили понимание филогении хитрида и привели к образованию нескольких новых зооспорических грибов :
- Отряд Blastocladiales , первоначально входивший в состав Chytridiomycota, теперь классифицируется как отдельный тип, Blastocladiomycota .
- Neocallimastigales , первоначально порядок анаэробных грибов класса хитридиомицет, найденный в пищеварительном тракте травоядных, позже был повышен до отдельного филы, то Neocallimastigomycota .
- Olpidiaceae , в том числе типа рода Olpidium , ранее классифицированном в порядке Chytridiales , были подняты в отдельный филюм, то Olpidiomycota .
Место обитания
Хитридиомикотики классифицируются в зависимости от положения, которое они занимают по отношению к субстрату: на субстрате (эпибиотики) или внутри субстрата (эндобиотики).
Они живут в почве в разлагающемся органическом веществе, на поверхности растений или животных, а также в воде. Они расположены от тропиков до Арктики.
Некоторые виды являются паразитами других грибов, таких как водоросли, планктон и сосудистые растения, а также животных.
В пресной воде они встречаются в ручьях, прудах и эстуариях. В морских экосистемах в основном паразитирует на компонентах водорослей и планктона. Виды Chytridiomycot встречаются от прибрежных отложений до глубины 10 000 м.
Возможно, большинство видов Chytridiomicota обитают в наземных средах обитания, таких как леса, сельскохозяйственные и пустынные почвы, а также в кислых болотах в качестве сапротрофов тугоплавких субстратов, таких как пыльца, хитин, кератин и целлюлоза в почве.
Некоторые виды обитают в тканях растений как облигатные паразиты; как в листьях, так и в стеблях и корнях.
Виды отряда Neocallimastigales обитают в рубце травоядных животных. Благодаря своей способности разрушать целлюлозу, они играют важную роль в метаболизме рубца.
Экологические функции Chytridiomycota
- Батрахохитриум Дендробатидис: Это хитрид Batrachochytrium, вызывающий хитридиомикоз. Это болезнь, поражающая земноводных. Это заболевание было обнаружено в Австралии и Панаме в 1998 году. Считается, что оно является основной причиной глобального сокращения численности амфибий. Этот гриб стал причиной исчезновения в 1989 году золотой жабы и гибели значительной части популяции жаб-брызгов киханси в Танзании.
- Другие паразиты: К другим паразитам относятся хитриды, которые в основном заражают водоросли, а также другие эукариотические или прокариотические микробы. Инфекция может нанести серьезный ущерб первичной продукции озера. Согласно некоторым сообщениям, паразитические хитриды могут оказывать значительное влияние на пищевые сети озер и прудов. Виды растений также могут быть заражены хитридами; Одним из примеров является Synchytrium endobioticum, ключевой патоген картофеля.
- Сапробы: Важнейшей экологической функцией, которую могут выполнять хитриды, является их разложение. Эти вездесущие, глобальные организмы ответственны за разложение тугоплавких веществ, таких как пыльца и целлюлоза, хитин и кератин. Хитриды также могут жить на пыльце и прикреплять к пыльцевым зернам нитевидные структуры, называемые ризоидами.
Экологические функции
Читридовые паразиты морских диатомовых водорослей. (А) Хитридовые спорангии на Pleurosigma sp. Белая стрелка указывает на поры сверхчувствительного отделяемого. (B) Ризоиды (белая стрелка) распространяются на хозяина диатомовой водоросли. (C) Агрегаты хлорофилла, локализованные в местах инфекции (белые стрелки). (D и E) Одиночные хозяева, несущие несколько зооспорангиев на разных стадиях развития. Белая стрелка на панели E выделяет ветвящиеся ризоиды. (F) Эндобиотические хитридоподобные спорангии в створке диатомовых водорослей. Штанги = 10 мкм.
Пеннатная диатомовая водоросль из Арктики плавильный пруд, инфицированные двумя хитридоподобными грибковыми патогенами спорангиев (выделены красным цветом).
Batrachochytrium dendrobatidis
Читрид Batrachochytrium dendrobatidis Ответственный за хитридиомикоз, болезнь амфибий. Обнаружен в 1998 г. Австралия и Панама эта болезнь, как известно, убивает амфибии в большом количестве и был предложен в качестве основной причины всемирного упадок амфибий. Было установлено, что вспышки грибка привели к гибели большей части Киханси Спрей Жаба популяция в естественной среде обитания Танзания, а также исчезновение золотая жаба в 1989 году. Хитридиомикоз также был причастен к предполагаемому исчезновению южной желудочной лягушки-выводка, в последний раз видели в дикой природе в 1981 году, а северную желудочную лягушку-выводок в последний раз встречали в дикой природе в марте 1985 года. Считается, что процесс, приводящий к гибели лягушек, заключается в потере основных ионов через поры, образованные хитридом в эпидермальных клетках во время его репликации.
Недавние исследования показали, что небольшое повышение уровня соли может вылечить хитридиомикоз у некоторых видов австралийских лягушек. хотя необходимы дальнейшие эксперименты.
Прочие паразиты
Хитриды поражают в основном водоросли и др. эукариотический и прокариотический микробы. Инфекция может быть настолько серьезной, что ограничивает первичную продукцию в озере. Было высказано предположение, что паразитические хитриды оказывают большое влияние на пищевые сети озер и прудов. Хитриды могут также поражать виды растений; особенно, Synchytrium endobioticum это важный картофель возбудитель.
Сапробы
Возможно, самая важная экологическая функция хитридов — разложение. Эти вездесущие и космополитические организмы ответственны за разложение тугоплавких материалов, таких как пыльца, целлюлоза, хитин, и кератин. Есть также хитриды, которые живут и растут на пыльце, прикрепляя нитевидные структуры, называемые ризоидами, на пыльцевые зерна. В основном это происходит во время бесполого размножения, потому что зооспоры, которые прикрепляются к пыльце, непрерывно воспроизводятся и образуют новые хитриды, которые будут прикрепляться к другим зернам пыльцы для получения питательных веществ. Эта колонизация пыльцы происходит весной, когда в водоемах накапливается пыльца, падающая с деревьев и растений.
Мертвая лягушка при хитридиомикозе (Б. dendrobatidis) приметы
Уникальная единая клетка
Отдел хитридиомикота представляет собой группу организмов, обладающих уникальной формой питания. В основе этой формы лежит единая клетка, которая объединяет в себе черты животного и гриба.
Эта клетка способна поглощать органические вещества, как животные, через фагоцитоз, и абсорбировать питательные вещества, как грибы, через эндоцитоз. Таким образом, она объединяет в себе лучшие черты обоих царств и способна вести более эффективный образ жизни.
Примером хитридиомикота с такой формой питания является род Choanoflagellida. Эти одноклеточные организмы имеют характерную форму колбы с одним или несколькими хвостовиками, окруженными флагелями. Они обитают в пресных и морских водоемах и питаются органическими веществами, которые поглощают с помощью фагоцитоза.
Суммируя, можно сказать, что уникальная единая клетка в отделе хитридиомикота представляет собой удивительное сочетание черт животного и гриба, что позволяет этой клетке обладать особыми способностями питания и адаптации к различным условиям окружающей среды.
Жизненные циклы паразитарных хитридиомикотиков
У паразитарных Chytriomycotes цикл обычно проще.
У паразитарных грибов растений, таких как Olpidium viciae, зигота подвижна и поражает растение-хозяин. Устойчивые споррангии развиваются внутри растительных клеток. Внутри этих спорангиев возникает кариогамия.



Наконец происходит прорастание и высвобождение зооспор. Зооспоры также могут напрямую инфицировать растение-хозяин.
В грибке Batrachochytrium dendrobatidis паразит на коже лягушки, после периода подвижности менее 24 часов зооспоры реабсорбируют свои жгутики и энцисты. Затем они прорастают в ризоиды, которые цепляются за кожу лягушки.
Слоевище развивается в спорангии, дающие начало новому поколению зооспор. Половая фаза пока неизвестна.
ссылки
- Alexopoulus, C.J., Mims, C.W. и Blackwell, M. Editors. (1996). Вступительная микология. 4го Нью-Йорк: Джон Вили и сыновья.
- Буссе Ф., Барткевич А., Терефе-Аяна Д., Нипольд Ф., Шлейснер Ю. и др. (2017). Геномные и транскриптомные ресурсы для разработки маркеров в Синхитриум эндобиотик, Неуловимый, но серьезный картофельный патоген. Фитопатологии. 107 (3): 322-328. doi: 10.1094 / PHYTO-05-16-0197-R
- Dighton, J. (2016). Экосистемные процессы грибов. 2й Бока Ратон: CRC Press.
- Кавана К. Редактор. (2017). Грибы: биология и применение. Нью-Йорк: Джон Уайли
- C., Dejean, T., Savard, K., Millery, A., Valentini, A. и др. (2017). Инвазивные североамериканские лягушки передают смертельный гриб Батрахохитрий дендробатидис инфекции местным видам хозяев-амфибий. Биологические инвазии. 18 (8): 2299-2308.
Половое размножение Chytridiomycota
У моноблеф оогамное половое размножение. В трех из шести родов моноблеф происходит слияние между неподвижным эмбрионом и подвижным оплодотворенным сперматозоидом. Сперматозоиды с одиночными жгутиками встречаются у Antheridia. Оосферы можно производить в Оогонии. Сперматозоиды оплодотворяют оосферы. Зигота образует толстую стенку, которая используется при многоплодии.
Зигота Monoblepharis становится подвижной после завершения оплодотворения. Жгутик сперматозоида приводит в движение зиготу. Однако зиготы Monoblepharella или Gonapodya сразу же образуют толстые стенки и после оплодотворения становятся ооспорами. Репродуктивный цикл регулируется предпочтительным температурным и световым режимами. Сюда относится бесполое размножение при более низких температурах (8-15°С) и половое при более высоких температурах (25-35°С).
Исследования Siphonaria variabilis, Polyphagus Euglenae и Polyphagus euglenae под световым микроскопом предоставили дополнительные доказательства того, что слиянию ядер предшествует образование покоящихся спор в хитридах. Есть много стратегий, которые можно использовать для рекомендации генетики, в том числе:
- Как и у Synchytrium, патогенные виды растений, Слияние подвижных гамет.
- Совокупление гаметангиев — это когда один гаметангий передает свою протоплазму другому, как описано Sporophlyctis.
- Гаметангиальный контакт — это когда содержимое одного гаметангиума перемещается через трубку конъюгации в другой гаметангий. Зиготические покоящиеся споры образуются, как у Zygorhizidium.
- Соматогамия, включающая слияние ризоидальных нитей, как у Chytriomyces variabilis и Siphonaria hyalinus.
Половая фаза
В сексуальной фазе путем слияния цитоплазм (плазмогамия) и последующего слияния ядер (кариогамия) планогамет образуется зигота. Здесь начинается диплоидная фаза цикла.
Зигота может принимать форму резистентной кисты или трансформироваться в диплоидное ценоцитарное слоевище (споротальное).
Спороталы вызывают резистентные споррангии, которые образуют гаплоидные зооспоры, замыкая цикл. Из споротелия также могут образовываться зооспорангии, которые продуцируют диплоидные зооспоры, которые энцистизируют и при прорастании образуют новые споротелии.
Половое размножение также может происходить путем оогамии, как в случае отряда Monoblefaridales. Здесь неподвижная женская гамета, расположенная в оогониуме, принимает подвижную флагеллированную мужскую гамету.
Библиография
- Барр, DJS 2001. Chytridiomycota. стр. 93-112. В: Микота. VIIA Систематика и эволюция. (ред. DJ McLaughlin, EG McLaughlin, PA Lemke) Springer-Verlag Berlin.
- Чемберс, Дж. 2003. Рибосомная ДНК, вторичная структура и филогенетические отношения среди Chytridiomycota. Кандидатская диссертация. Университет Алабамы. 116 стр. август 2003 г.
- James, TY, D. Porter, CA Leander, R. Vilgalis, and JE Longcore 2000. Молекулярная филогенетика Chytridiomycota подтверждает полезность ультраструктурных данных в систематике хитрид. Канада. Дж. Бот. 78: 336-350.
- Летчер, П.М. 2003. Систематический анализ молекулярных и ультраструктурных признаков двух групп зооспоровых грибов. Кандидатская диссертация. Университет Алабамы. 149 стр. декабрь 2003 г.
- Longcore, JE, AP Pessier и KD Nichols. 1999. Batrachochytrium denderobatidis gen. и др. nov., хитрид, патогенный для амфибий. Микология 91: 219-227.
- Пауэлл, MJ 1993. Глядя на микологию с лицом Януса. Взгляд на хитридиомицеты в окружающей среде. Микология 85: 1-20.
- Воробей, Ф.К. 1960. Водные фикомицеты. 2-е издание ред. Мичиганского университета, Анн-Арбор, Мичиган