

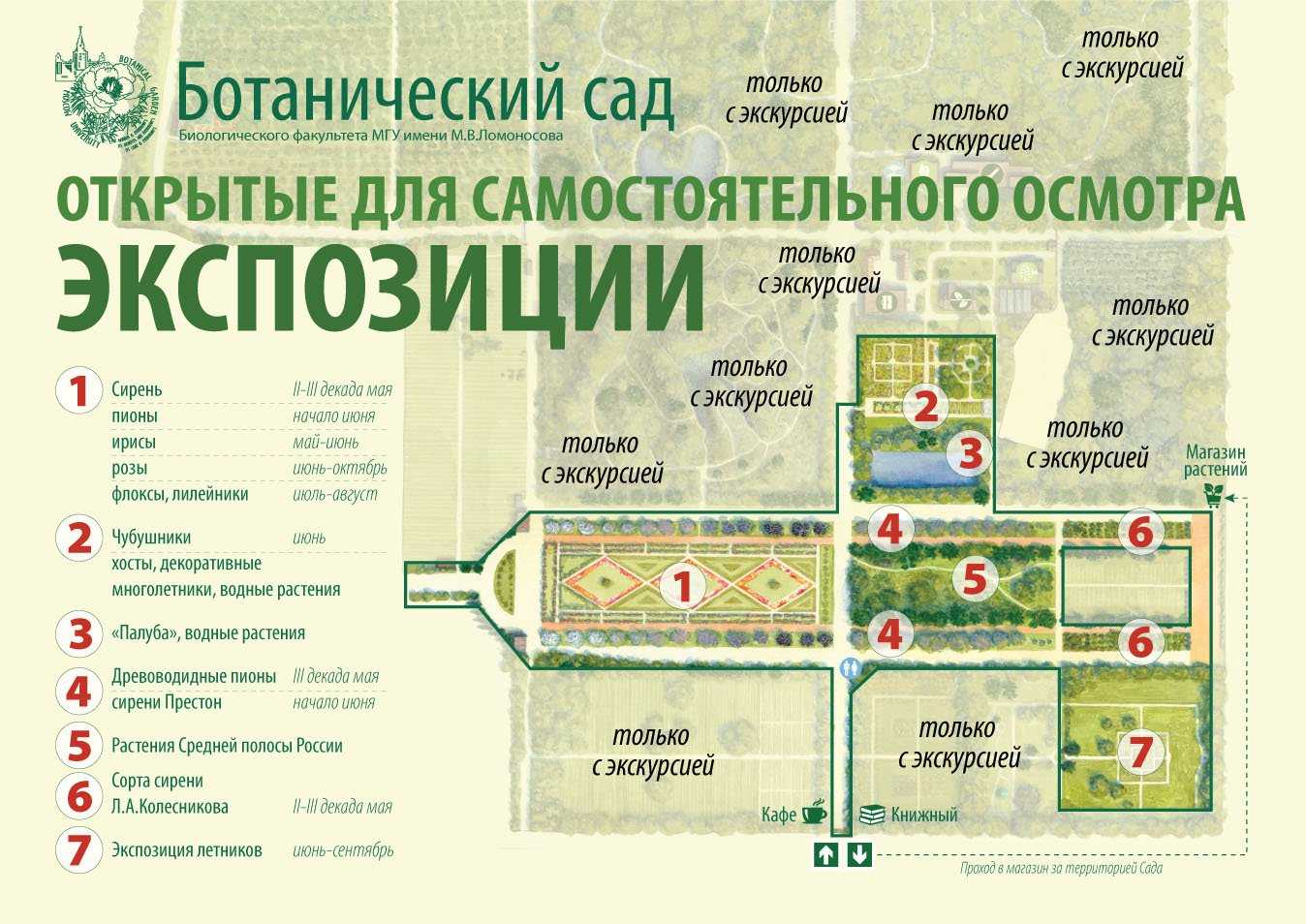
Лист
Типичный лист лиственных покрытосеменных состоит преимущественно из первичных тканей. Пластинка листа обычно широкая и плоская, поддерживается черешком, содержит основную ткань, или мезофилл, окруженный сверху и снизу эпидермой.
Питательные вещества, вода и минеральные соли перемещаются и листьях по жилкам, густо пронизывающим мезофилл. Ткань мезофилла содержит многочисленные межклеточные пространства, сообщающиеся с внешней средой отверстиями (устьицами) в эпидерме. Устьица играют существенную роль в физиологии растений, так как они служат воротами, через которые испаряется вода из листьев и поступает углекислый газ, используемый в фотосинтезе. У большинства лиственных деревьев устьица располагаются только на нижней стороне листа.
Листья образуются за счет деления поверхностных клеток апикальной меристемы. Появляющиеся листья располагаются очень тесно друг к другу, узлы и междоузлия почти неразличимы. Затем, когда активируется меристема между листьями, начинается интеркалярный рост и тогда обозначаются междоузлия. Известно несколько типов образования листьев кроны. У некоторых видов максимальная площадь листьев достигается уже в начале вегетационного периода и на протяжении года новые листья не появляются. У других видов новые листья в течение сезона добавляются за счет постоянного образования и разрастания новых листовых зачатков или путем нескольких периодических вспышек роста.
«ВОЛНОВОЕ» ОБРАЗОВАНИЕ ПОБЕГОВ. У многих деревьев рост побегов происходит периодически в виде волн или вспышек в течение вегетационного периода. При этом на побеге распускается несколько терминальных почек в год. После формирования побега из первой почки с ее фиксированными дополнительными листьями сразу же образуется вторая почка на апексе того же самого побега. Эта почка очень скоро распускается, давая удлиненный побег. За второй фазой роста могут следовать дополнительные волны роста из почек, последовательно распускающихся возле верхушки того же осевого побега. У цитрусовых, в зависимости от локализации побегов на дереве и климатических условий, обычно имеется две мощные и до трех слабых волн побегообразования в год. У молодых деревьев периоды побегообразования бывают чаще, чем у старых деревьев.
Продолжительность разворачивания отдельных листьев у разных видов различна. Листья вечнозеленых покрытосеменных обычно разворачиваются медленно. Листья цитрусовых развиваются до 130 дней. Даже после того, как листовые пластины полностью развернутся, они продолжают утолщаться и у них увеличивается сухой вес.
Не все почки образуют побеги. Некоторые из них остаются спящими на протяжении всей жизни дерева. Спящие почки, заложенные в пазухах листьев, соединяются с корой через листовой след. Побегообразование из спящих почек — обычное явление. Почки, которые формируются на более старых частях растения, а не на верхушках стебля или в пазухах листьев, называются адвентивными. Они образуются на частях корня или ствола, у которых нет связи с апикальной меристемой. Они могут возникать как из глубоко расположенных, так и из поверхностных тканей. Например, адвентивные почки могут возникнуть из тканей каллюса вокруг раны, из камбия или из зрелых тканей эндодермы.
МАТЕРИАЛЫ И МЕТОДЫ
Сборы генеративных особей проводились в конце июня в окрестностях с. Усть-Кан (Горный
Алтай) в двух ценопопуляциях (ЦП 1, 2) и в лесопарковой зоне Центрального сибирского
ботанического сада (ЦСБС) (г. Новосибирск) (ЦП 3). Для изучения внутрипочечного развития
сбор материала проводился только в ЦП 3 в несколько сроков: в конце апреля, начале
июня, конце июля и в начале октября. В каждом местообитании были изучены по 30–45
цветущих особей. Для оценки мощности развития измерялись длина генеративного побега
и его паракладиев, определялась масса побега и масса частей побега, несущих генеративные
органы (на главной оси и на паракладиях) и число цветков в одном пазушном дихазии
на главном соцветии и репродуктивное усилие особи. Все полученные данные статистически
обработаны в пакете программ Exсel. Достоверность различий между параметрами определялось
по по t-критерию Стьюдента при 5%-м уровне значимости (Zaitsev, 1991).
ЦП 1 входит в состав злаково-разнотравной луговой степи на опушке лиственничника по северо-западному
склону (уклон 5°). Общее покрытие (ОПП) – 70%. Проективное покрытие (ПП) P. tuberosa – 8%. ПП злаков составляет 10%: Poa angustifolia (5%), Koeleria cristata (3%), Poa attenuata (3%). Разнотравье представлено Potentilla chrysantha (5%), Galatella biflora (5%), Ligularia glauca (5%), Fragaria viridis (5%), Adonis vernalis (3%) и др. ЦП 2 расположена в нижней части юго-восточного склона (крутизна 30°) в составе каменистой
закустаренной луговой степи. ОПП составляет 80%. ПП P. tuberosa – 3–5%. Из кустарников присутствуют Spirea trilobata (8%), Caragana pygmaea (1%). ПП злаков составляет 15%: Phleum phleoides (5%), Poa botrioides (5%), Koeleria cristata (3%), Agropyron pectinatum (3%). Из разнотравья произрастают: Astragalus austriacus (5%), Onobrychis arenaria (3%), Artemisia frigida (3%), Artemisia commutata (3%), Bupleurum bicaule, B. multinerve, Dianthus versicolor, Thymus mongolicus и др. ЦП 3 входит в состав разнотравно-злакового луга, расположенного на северо-западном склоне
(уклон 12°). ОПП травостоя – 80%. ПП P. tuberosa – 2%. ПП злаков – 60%, в том числе Dactylis glomerata (30%), Festuca pratensis (20%), Phleum pretense (5%), Poa angustifolia (5%). Из разнотравья присутствуют Fragaria viridis (20%), Lathyrus pratensis (10%), Heiracium umbellatum (5%), Trifolium pratense (5%), Vicia amoena (5%), Viola hirta (2%) и др. виды. Заметное покрытие имеют индикаторы антропогенного воздействия: Cirsium setosum (10%), Plantago major (3%) и Urtica dioica (3%).
Функциональное зонирование побеговой системы определялось согласно W. Troll (1964),
И.В. Борисовой, Т.А. Поповой (Borisova, Popova, 1990), тип соцветия – по Т.В. Кузнецовой
и А.К. Тимонину (Kuznetsova, Timonin, 2017). Морфогенез побега описан по представлениям
Т.И. Серебряковой (Serebryakovа, 1971). Репродуктивное усилие вычислялось как доля
веса репродуктивных органов от общего веса особи в процентах (Markov, Pleshchinskaya,
1987).
Сибирский экологический журнал
2020 год, номер 3
Модель побегообразования как основа адаптаций цветковых растенийН.П. Савиных, С.В. Шабалкина Вятский государственный университет, Киров, Russiasavva_09@mail.ru Аннотация Работа направлена на описание биоморфологии растений, обитающих в условиях переменного обводнения. Полученные результаты расширяют представления о концепции модульной организации как конструктивного и репрезентативного способа сравнительно-морфологического анализа и выяснения механизмов эволюции биоморф. С позиций системного и комплементарного подходов изучена структурная организация биоморф в градиенте изменения устойчивости увлажнения в направлении пойменные луга — прибрежья — урез воды и пляжи на примере модельных видов: Solanum dulcamara , Thalictrum simplex , Lysimachia vulgaris , Lycopus europaeus , Stellaria palustris , Persicaria amphibia , Erysimum cheiranthoides . Установлено, что акселерация в вегетативной и генеративной сферах приводит к возникновению монокарпиков в гигрофильной линии эволюции. Показано, что онтогенез особей семенного и вегетативного происхождения сокращается до единственного плодоношения, а специфика модели побегообразования сохраняется у растений во всех рассмотренных биотопах. Из специфических признаков выявлены: наличие длинных междоузлий у побегов в субстрате, в надсубстратной части при подтоплении, в травяной ветоши и густом травостое при низкой освещенности; симподиальное нарастание путем образования побегов замещения. На модельных видах определено, что приспособленность растений обеспечивают изменения отдельных структурных элементов монокарпических побегов, а структурно-функциональная зональность монокарпических побегов обусловливает автономизацию частей особи. Наши результаты позволили подтвердить, что монокарпический побег и система, сформировавшаяся на его основе, — одна из категорий осей травянистого растения в составе архитектурной единицы (в понимании D. Barthelemy и Y. Caraglio). Полученные данные показали, что заселение водоемов цветковыми растениями возможно от длиннопобеговых длиннокорневищных трав и полукустарниковых лиан в следующих направлениях: 1) возникновение монокарпиков однолетников; 2) возникновение малолетников и однолетников вегетативного происхождения с формированием ацентрических и явнополицентрических столонообразующих и корнеотпрысковых биоморф. |
РОСТ ПЛОДОВ.
Период от цветения до созревания плодов продолжается от 3 недель (у земляники) до 60 недель (у некоторых сортов апельсина). Различные плоды растут с разной скоростью и достигают в зрелом состоянии различных размеров. Например, маслины растут очень медленно (около 0,01-0,02 см^ в день). Следует отметить, что скорость роста плодов значительно зависит от сезона, условий и агротехнических приемов. Различна она также у разных плодов одного сбора
Рост плодов происходит за счет клеточных делений и увеличения размеров клеток. В период цветения клеточное деление ограничено, но после завязывания плодов многие их ткани становятся меристематическими, к ним поступает очень много углеводов.
![Устройство газонов [1987 ерохина в.и., жеребцова г.п., вольфтруб т.и., покалов о.н., шурова г.в. - озеленение населенных мест. справочник]](https://lesniepolyani.ru/wp-content/uploads/2/9/b/29bc8ff03378031cc120f0f19c83ea3b.jpeg)








У некоторых плодов (например, у смородины, ежевики) деление клеток (за исключением клеток зародыша и эндосперма) заканчивается к моменту опыления. У яблони и цитрусов клетки делятся в течение короткого времени после опыления. А у авокадо клетки продолжают делиться в течение всей жизни плода. У большинства видов увеличение размеров клеток делает основной вклад в общий рост плода. У винограда, например, за счет увеличения количества клеток размер плода удваивается, а за счет увеличения объема клеток размер плода возрастает в 300 раз.
Деревья всегда рассматривались не-ботаниками как представители вершины развития растительного царства. Из-за размеров и красоты, их часто наделяли особыми свойствами и даже поклонялись им. У деревьев действительно есть специфические свойства, но эти отличия носят скорее количественный характер, чем являются отличиями по существу от других растений. Деревья проходят те же стадии роста, им присущи те же процессы, как и другим семенным растениям Только их большие размеры, медленное развитие и продолжительность жизненного цикла придают им некоторые особенности, отличные от тех, которые присущи небольшим растениям с более коротким жизненным циклом.
Для выращивания деревьев необходимо понимать основные процессы, управляющие их ростом и развитием, а также влияние факторов окружающей среды и агротехнических мероприятий. Таким образом, знание физиологии растений очень способствует достижению успехов на садоводческом поприще.
Подробнее — в книге «Физиология древесных растений» (П.Крамер, Т.Козловский) см.Библиотека
СТВОЛ
Ствол древесных растений поддерживает крону, проводит воду и минеральные вещества вверх от корня, передает питательные вещества и гормоны от места их синтеза к участкам, где они используются в ростовых процессах или откладываются в запас. На верхушке ствола и на каждой ветке имеется терминальная точка роста, определяющая увеличение в длину. Между корой и древесиной ствола, ветвей и главного корня располагаются камбий и тонкий слой покровной латеральной меристемы.
Ранения и заживление ран. Раны на деревьях служат путями для патогенных организмов. Раны возникают в результате поломки ветвей, механических повреждений, ранений животными, пожарами и т.д. Скорость и эффективность реакции растения на раневое воздействие зависит, с одной стороны, от глубины и степени повреждения, с другой стороны, от состояния растения. Раны, повредившие только кору или слегка задевающие камбий, обычно заживают быстро.
Процесс заживления заключается в затягивании ран и изолировании прилегающих к ранам тканей, пораженных микроорганизмами. Многие обширные раны старых деревьев никогда не затягиваются, но заживляются.
Заживление глубоких ран ствола происходит благодаря образованию каллюса и формированию нового камбия путем превращения клеток каллюса в камбиальные клетки. Меристема также регенерирует в процессе заживления ран. Образование каллюса у разных видов происходит с разной скоростью. Формирование нового камбия не зависит от количества или скорости образования каллюса.
КОРНИ
 Корни служат для закрепления растения в почве, поглощения воды и минеральных солей, а также для запасов питательных веществ. Корневая система деревьев состоит из сети относительно крупных многолетних корней и множества небольших недолговечных боковых корней. Размер корневой системы сильно зависит от типа почвы. Корни плодовых деревьев на песчаных почвах распространяются в 3 раза дальше пределов кроны, на суглинистых почвах — в 2 раза, на глинистых — только в 1,5 раза.
Корни служат для закрепления растения в почве, поглощения воды и минеральных солей, а также для запасов питательных веществ. Корневая система деревьев состоит из сети относительно крупных многолетних корней и множества небольших недолговечных боковых корней. Размер корневой системы сильно зависит от типа почвы. Корни плодовых деревьев на песчаных почвах распространяются в 3 раза дальше пределов кроны, на суглинистых почвах — в 2 раза, на глинистых — только в 1,5 раза.
Длинные корни функционируют на протяжении всей жизни дерева и увеличиваются в диаметре за счет активности камбия. Короткие корни, располагающиеся по бокам длинных корней, имеют апикальную меристему и лишены истинного корневого чехлика. Короткие корни лишены вторичного роста и большая их часть исчезает в первые год-два жизни, заменяясь новыми.
Общие сведения
На различных этапах развития человеческого общества деревья для людей имели разное значение. Для наших далеких предков они были источником топлива и убежищем, а иногда и предметом поклонения. Для горожан они создавали летом приятную тень, хотя осенью и причиняли неудобства, засыпая листьями газоны. Для работников парков деревья являются декоративными объектами ландшафта.


Деревья служат источником сырья для бумажной и деревообрабатывающей промышленности. Садоводы изучают деревья как источник плодов. Для физиологов деревья являются сложными биохимическими фабриками, которые буквально строят сами себя. Поэтому физиологи изучают многочисленные процессы, которые в совокупности обеспечивают то, что мы называем ростом.







![Устройство газонов [1987 ерохина в.и., жеребцова г.п., вольфтруб т.и., покалов о.н., шурова г.в. - озеленение населенных мест. справочник]](https://lesniepolyani.ru/wp-content/uploads/b/7/e/b7e60682e7ede24597f4f6b074e84ccf.jpeg)

Неблагоприятные окружающие условия, влияющие на физиологические процессы, снижают рост дерева. Например, водный дефицит тормозит рост, так как при этом закрываются устьица, снижается интенсивность фотосинтеза, прекращается рост клеток и возникают другие неблагоприятные условия внутри дерева. Снижает рост дерева и недостаток азота, который является существенной составной частью белков, необходимых для формирования новой протоплазмы, ферментов и других необходимых веществ. Очень важны для роста дерева фосфор, калий, кальций, сера и другие минеральные элементы, входящие в состав коферментов, буферных и других биохимических систем, необходимых для осуществления различных биохимических процессов.
НАИБОЛЕЕ ВАЖНЫЕ ФИЗИОЛОГИЧЕСКИЕ ПРОЦЕССЫ
Фотосинтез — синтез углеводов из углекислого газа и воды в хлорофиллоносной ткани деревьев. Углеводы — основные питательные материалы, используемые в других процессах.
Азотный обмен — включение неорганического азота в органические соединения, что делает возможным синтез белка и протоплазмы.
Липидный, или жировой, обмен — синтез липидов и родственных им соединений.
Дыхание — окисление питательных веществ в живых клетках, в результате которого высвобождается энергия, используемая при ассимиляции, поглощении минеральных веществ и других процессах, идущих с затратами энергии.
Ассимиляция — преобразование питательных элементов в различные структуры; процесс роста.
Аккумуляция питательных элементов — запасание питательных веществ в семенах и паренхимных клетках древесины и коры.
Абсорбция — поглощение’ воды и минеральных веществ из почвы, кислорода и углекислого газа из воздуха.
Транслокация — передвижение воды, минеральных и питательных веществ и гормонов от одной части дерева к другой.
Транспирация — потеря воды в виде паров.
Рост — постоянное увеличение размеров в результате взаимодействия физиологических, перечисленных выше процессов.
Генеративное размножение — образование в результате взаимодействия многих физиологических процессов цветков, плодов и семян.
Вегетативное размножение — играет важную роль у некоторых видов. У многих древесных растений с возрастом способность к вегетативному размножению снижается, что может представлять некоторую проблему при выращивании плодовых деревьев.







СТРОЕНИЕ ДЕРЕВА
Можно считать, что дерево состоит из шести частей.
Три части — листья, ствол, корни — вегетативные структуры;
и еще три части — цветы, плоды и семена — репродуктивные органы.
Каждая из этих частей состоит из большого количества различных тканей.