

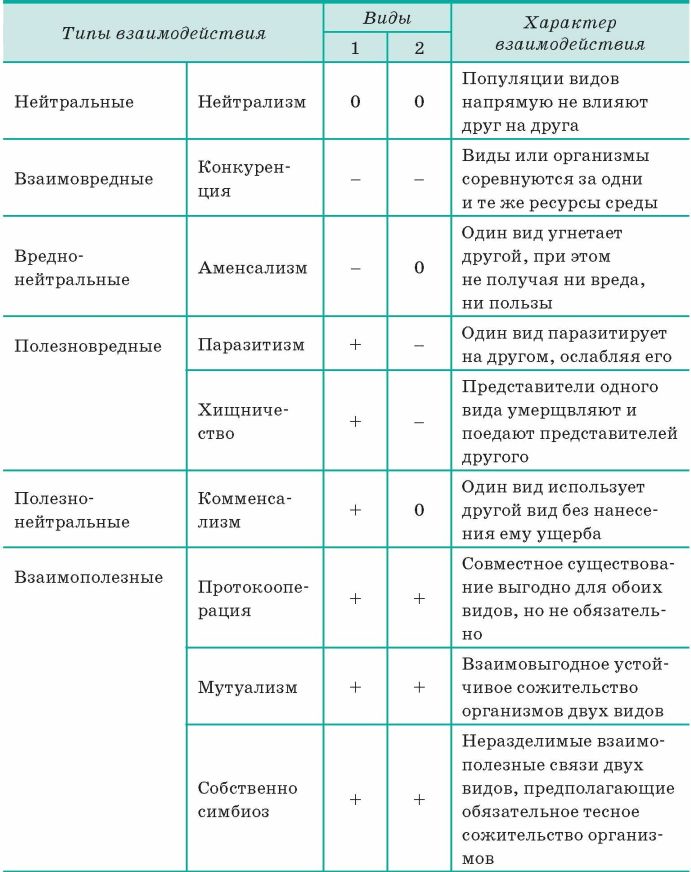
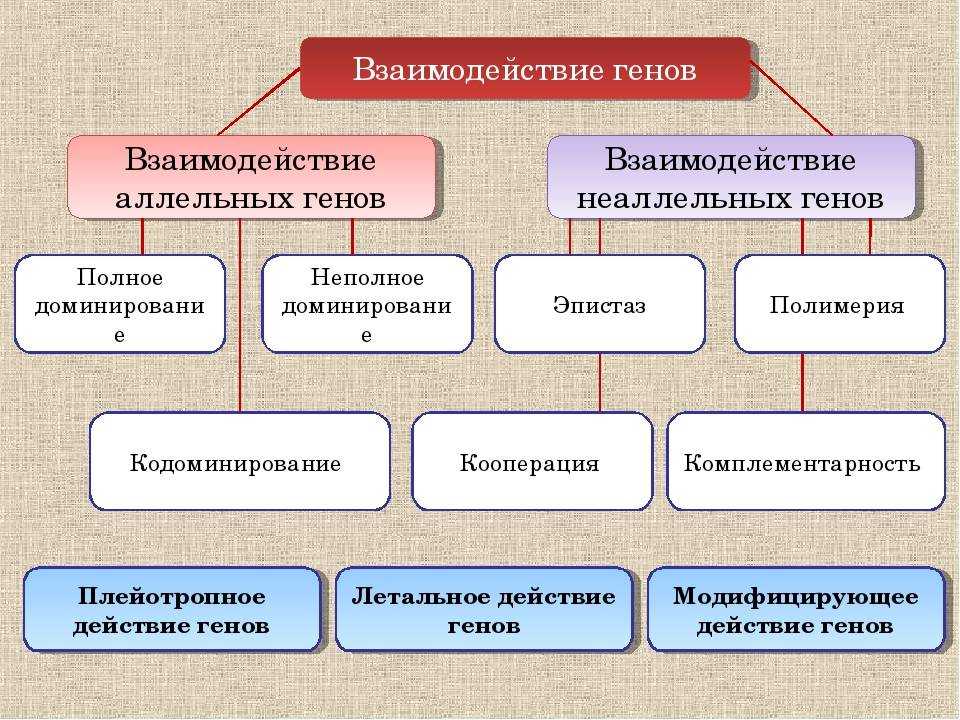
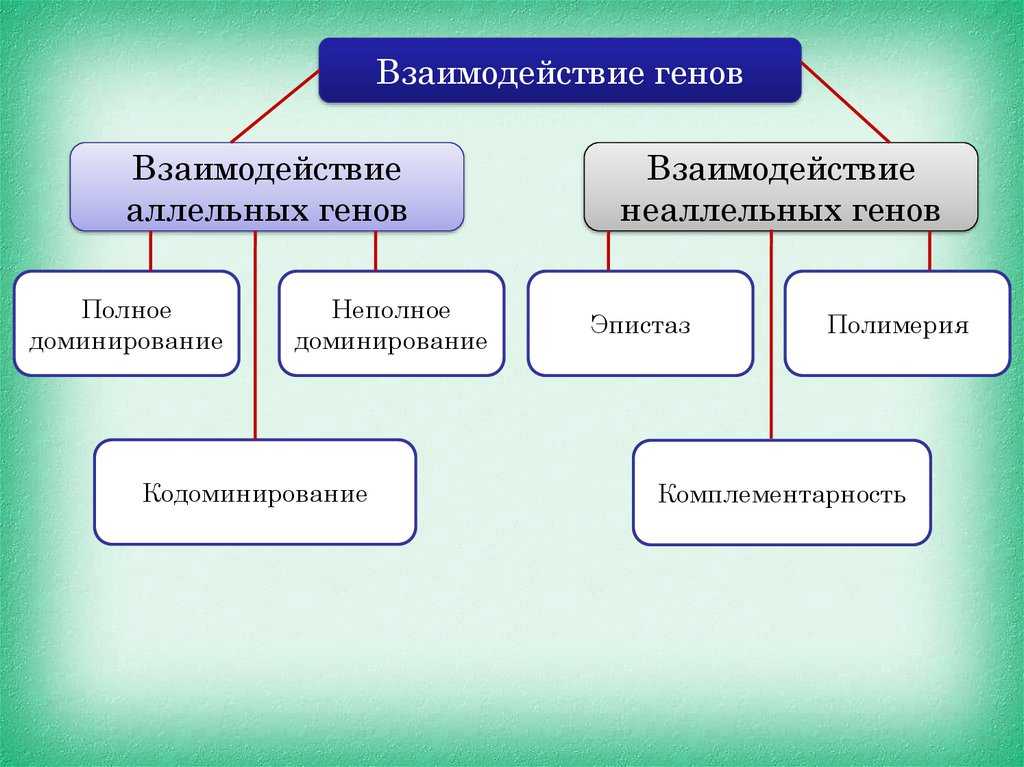
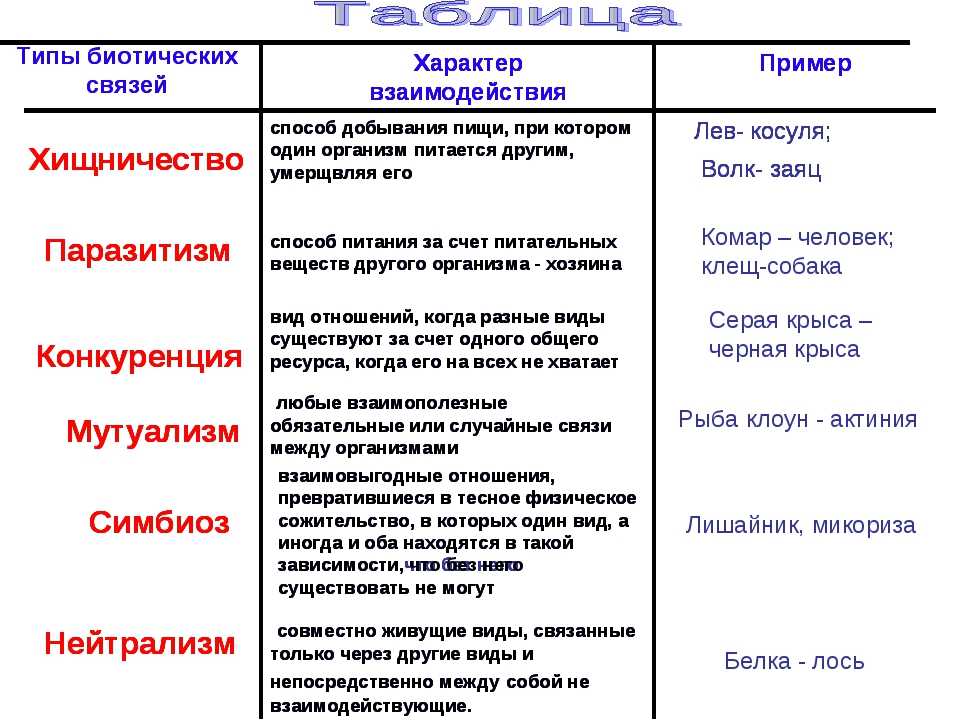
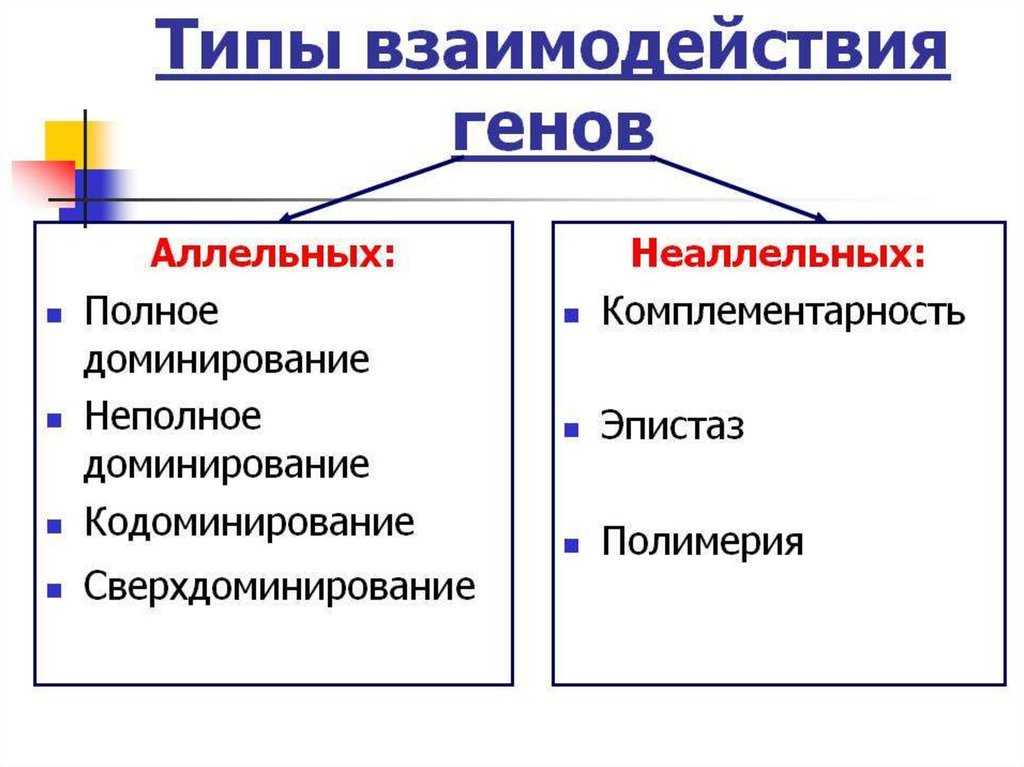
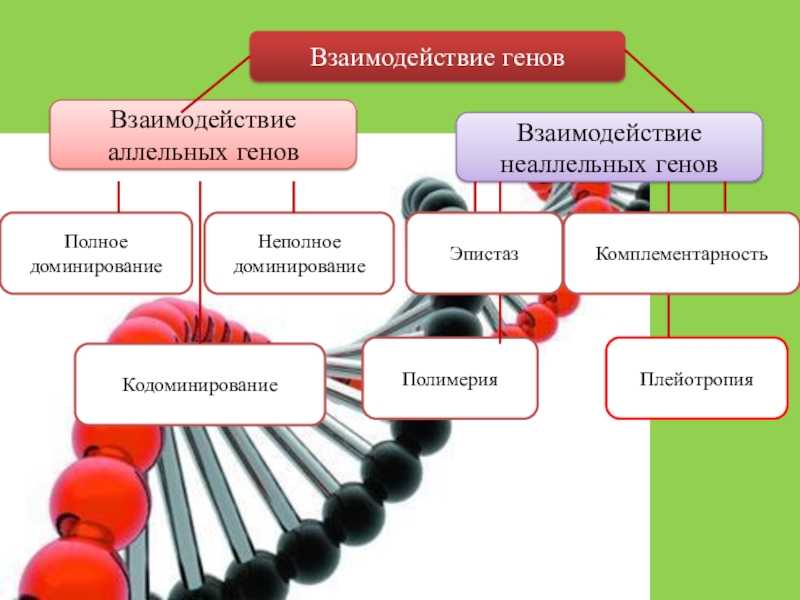
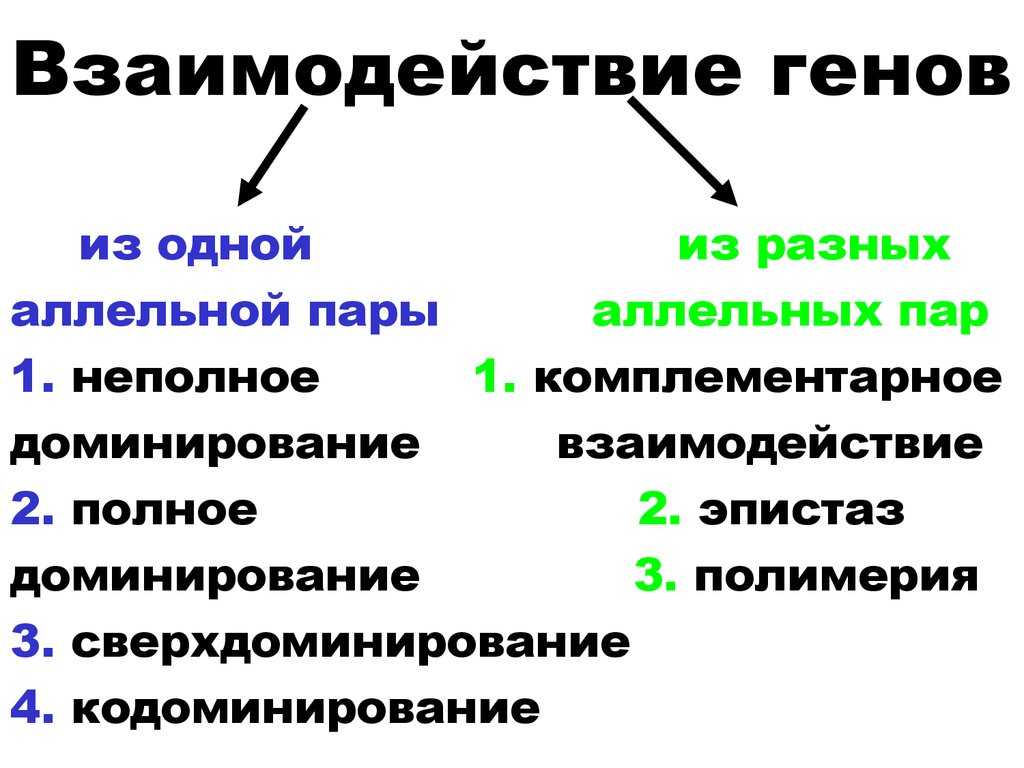
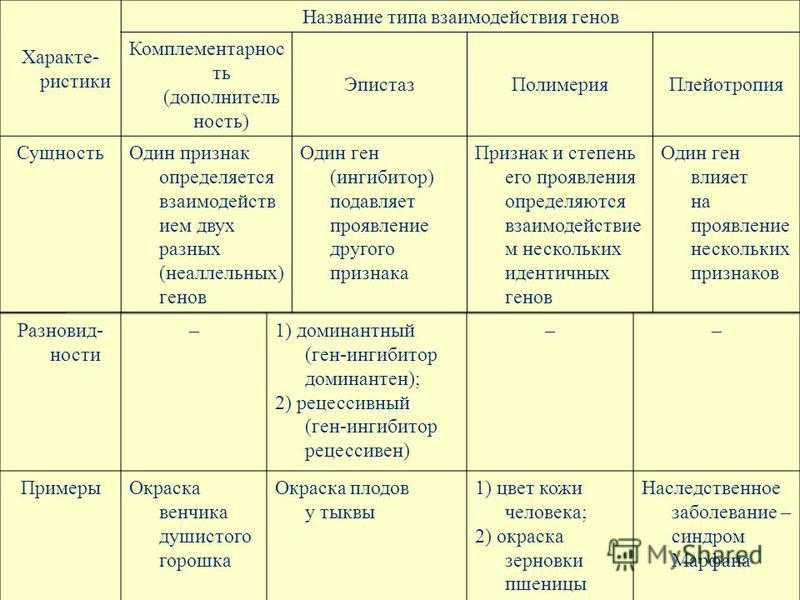
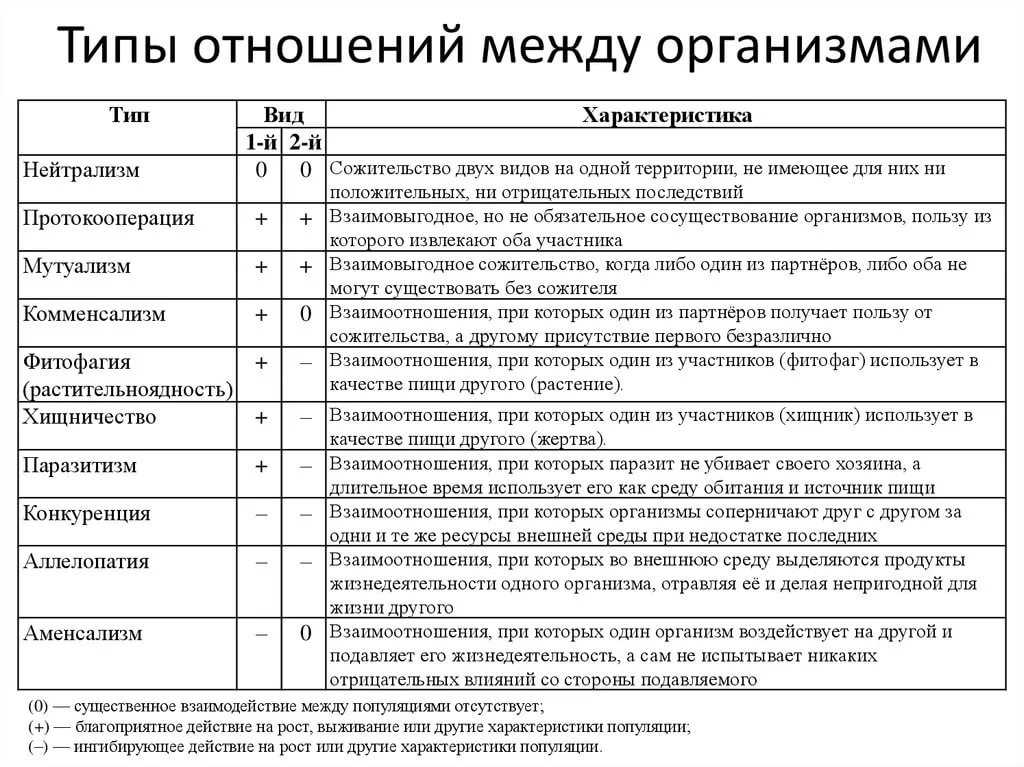
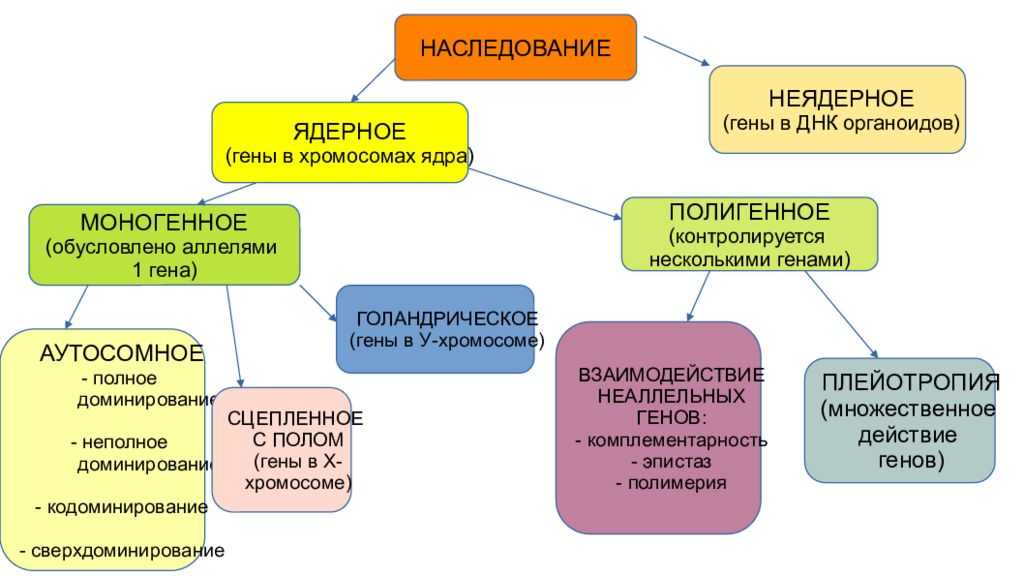
Взаимодействие аллельных генов
Аллели одного гена, находясь в одном организме, взаимодействуют между собой, и от этого взаимодействия зависит, как проявит себя признак, обусловленный соответствующим геном.
Полное доминирование
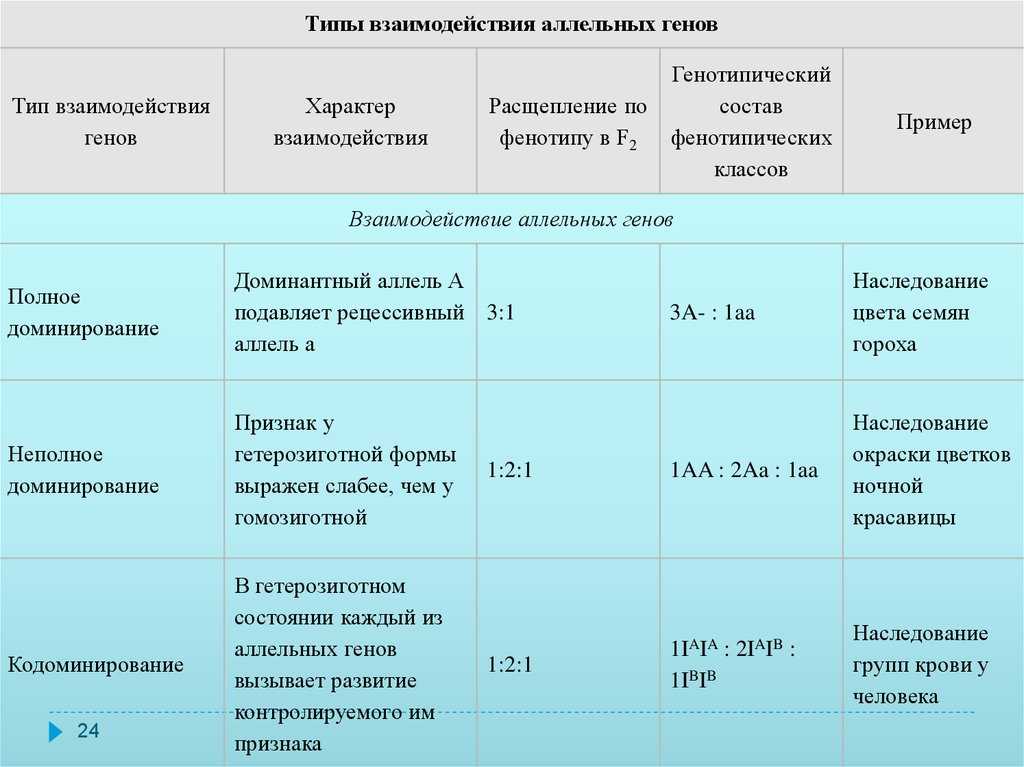
Чаще всего встречается такой тип взаимодействия как полное доминирование, при котором один аллельный ген проявляется и полностью подавляет проявление другого аллельного гена. В данном случае первый называется доминантным, а второй — рецессивным.
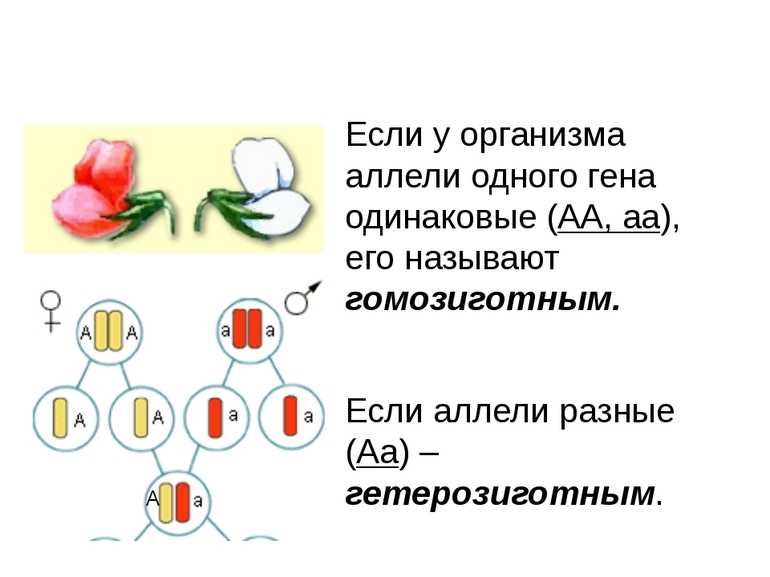
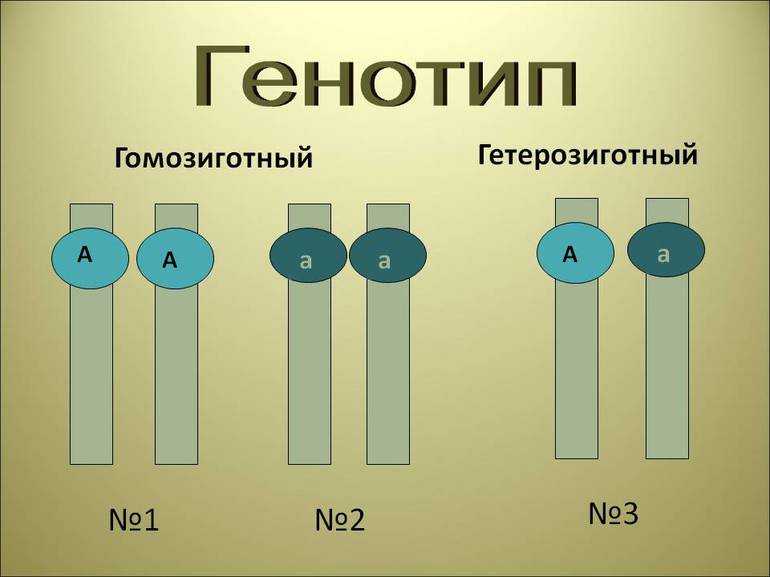
В генетике принято доминантный ген обозначать большой латинской буквой (например, ), а рецессивный — малой (). Если особь гетерозиготна, то ее генотип будет . Если гомозиготна, то или .
В случае полного доминирования генотипы и будут иметь одинаковый фенотип.
Полное доминирование наблюдается при проявлении, например, цвета семян гороха. У гетерозиготных () растений по этому признаку семена всегда желтые, как и у гомозиготных по доминантному аллелю (). Зеленый цвет определяется рецессивным аллелем и поэтому подавляется.
При множественном аллелизме отношения между аллелями могут быть неоднозначными. Если имеет место быть исключительно полное доминирование, то один ген может быть доминантным по отношению ко второму, но рецессивным по отношению к третьему. В таком случае строят ряды ( > > > …), в которых отражают отношения доминирования. Так, например, наследуется окраска шерсти у многих животных, цвет глаз.
Кроме полного доминирования встречаются и другие типы взаимодействия аллелей: неполное доминирование, кодоминирование, сверхдоминирование, комплементация аллелей и некоторые другие.
Неполное доминирование
В случае неполного доминирования гетерозиготный генотип будет иметь промежуточное значение признака. В этом случае доминантный аллель лишь частично подавляет рецессивный. В результате в фенотипе проявляется некое промежуточное значение признака.
Например, цвет цветков ночной красавицы с генотипом красный, — белый, — розовый.
При неполном доминировании значение признака не обязательно будет ровно «по середине». Чаще бывает отклонение в сторону доминантного, то есть когда фенотип генотипа хоть и не сходен с , но ближе к нему, чем к фенотипу генотипа .
Кодоминирование
При кодоминировании два аллельных гена, оказавшись в одном организме, полностью проявляют себя. Ни один из них никак не подавляет другой. В случае кодоминирования отсутствуют доминантно-рецессивные отношения между аллелями. В итоге в организме синтезируется два разных белка, определяющих один и тот же признак. Нечто похожее происходит при сверхдоминировании и межаллельной комплиментации.
При множественном аллелизме в одной паре аллелей может быть отношение полного доминирования, а в другой — кодоминирования. Так, например, обстоит дело с группами крови человека. Аллель является рецессивным и подавляется доминантными аллелями и . Если организм , то у него первая группа крови. Если или — вторая, или — третья. Если же человек имеет аллели и , то есть генотип , то у него четвертая группа крови, при которой в крови присутствуют два антигена (за наличие одного отвечает аллель , за наличие другого — ).
Сверхдоминирование
Сверхдоминирование — это такой тип взаимодействия аллельных генов, когда у гетерозиготных особей () признак сильнее проявляется, чем у обеих гомозиготных ( и ). Причина подобного явления лежит в достаточно сложных биохимических процессах, связанных с тем, что аллели кодируют несколько разные, но аналогичные по своим функциям полипептиды. В итоге признак как бы накапливается. Сверхдоминирование лежит в основе такого явления как гетерозис, когда дочерние организмы более жизнеспособны, чем родительские.
Помимо перечисленных существуют и другие более редкие типы взаимодействия аллелей.
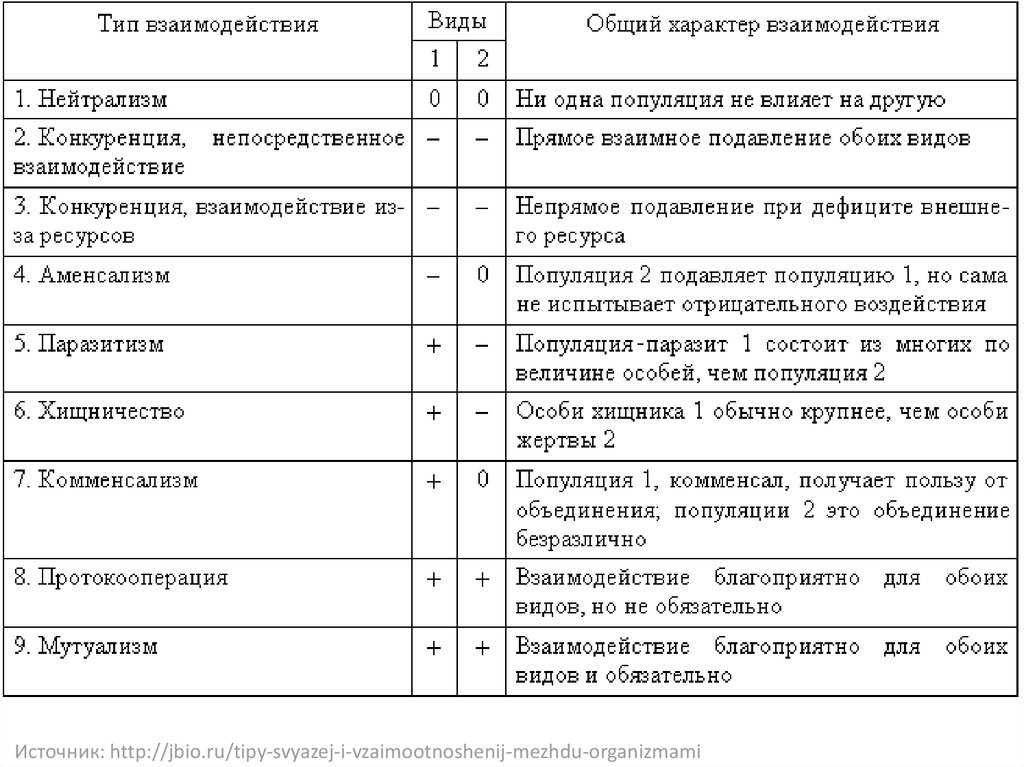
Неаллельные взаимодействия генов
Доминирование и рецессивность
– типичные примеры взаимодействия аллельных генов.
Однако в процессе индивидуального развития организма и неаллельные гены
вступают в сложные взаимодействия между собой. Организм – не мозаика,
складывающаяся из действия отдельных и независимых генов, а сложная
система последовательных биохимических и морфологических процессов,
определяемых совокупностью генов – генотипом
Понятие наследование признака употребляют обычно как образное
выражение. В действительности наследуются не признаки, а гены. Признаки
формируются в ходе индивидуального развития организма, которые
обусловливаются генотипом и влиянием внешней среды.
Принято различать следующие основные типы взаимодействия неаллельных
генов:
- Комплементарность,
- Эпистаз
- Полимерия.
Эпистаз.
Эпистатическое взаимодействие неаллельных генов в определенном
смысле противоположно комплементарному действию генов. Сущность
эпистаза сводится к подавлению проявления генов одной аллельной пары
генами другой. Гены, подавляющие действие других неаллельных генов,
называются супрессорами или подавителями. Они могут быть как
доминантными, так и рецессивными, например А — В- или bbA — .
Наследование окраски у свиней демонстрирует доминантный Эпистаз. При
скрещивании черных и белых свиней из разных пород в F1 появляются белые
потомки. Их скрещивание между собой приводит к появлению белых (12/16),
черных (3/16) и красных (1/16) поросят. Все белые поросята имеют
минимум один доминантный генподавитель I. Черные поросята гомозиготны
по рецессивному аллелю i, не препятствующему формированию окраски, и
несут доминантный аллель Е, детерминирующий образование черного
пигмента. Красные поросята (eeii) лишены доминантного геноподавителя I
и доминантного гена, определяющего черную окраску.
Полимерия.
| В некоторых случаях установлено, что проявление конкретного признака зависит от количества доминантных генов, вносящих вклад в его развитие. Например, при скрещивании краснозерных пшениц с белозерными было установлено, что растения с генотипом А1А1А2А2 имеют красные зерна, растения а1а1а2а2 – белые зерна, растения с тремя доминантными генами – красноватую окраску, а растения с двумя и одним геном – более бледную окраску. Таким образом, накопление определенных аллелей в генотипе может вести к изменению выраженности признаков. |
Кодоминирование и множественный аллелизм
Аналогичные расщепления характерны и для кодоминирования с той разницей, что потомки от скрещивания гомозигот разных типов будут характеризоваться наличием фенотипических признаков обоих родителей одновременно. То есть в этом случае оба аллеля являются доминантными: образуют функциональные продукты, которые немного отличаются по некоторым характеристикам. Такая ситуация довольно часто наблюдается для ферментов, когда два аллеля обуславливают синтез двух белков — изоферментов — с практически одинаковой активностью, но при этом они отличаются молекулярной массой (один из белков имеет дополнительный структурный домен и так далее).
Эти примеры требуют важного уточнения. Поскольку в двух гомологичных хромосомах могут находиться только два разных аллеля одного гена, ещё не означает, что в группе особей ген обязательно имеет два аллеля
На самом деле, в большинстве случаев гены существуют в виде нескольких (теоретически неограниченное количество) разных аллелей. Это явление получило название множественный аллелизм.
https://youtube.com/watch?v=FHkFuzgLBm0
Одним из примеров множественного аллелизма является система групп крови АВ0. Хорошо известны четыре группы крови — 0, А, В и АВ — определяются тремя аллелями одного гена: ІА, ІВ, і0. Аллели ІА и ІВ являются кодоминантными (отвечают за формирование на поверхности эритроцитов антигенов двух типов — А и В), но при этом доминируют над аллелем і0, который не продуцирует ни одного антигена.
Полное доминирование
Одним из главных понятий в генетике является понятие признака — некоторой определённой особенности организма или его части, по которой особь отличается от другой. Признаком в генетическом понимании можно считать любую характеристику на уровне внешних черт, физиологических или биохимических особенностей:
- вес;
- цвет;
- рост;
- набор веществ определённого типа в органах и клетках;
- совокупность определённых белков и так далее.
Все признаки одного организма называются фенотипом. Мендель осознал, что все характеристики, которые можно использовать в генетическом анализе, должны выявляться постоянно (независимо от возраста, внешних условий и другого) и должны иметь два или более альтернативных проявлений.
Аллельные гены (аллели) — это гены, отвечающие за один признак, но в разных его проявлениях (например, цвет семян — это один признак, который кодирует один ген, но проявляться может или жёлтый, или зелёный в зависимости от того, какой аллель является доминирующим).
Скрещивание, в котором проводится анализ одной пары альтернативных характеристик, называется моногибридным. Например, цвет семян гороха определяется геном sgr. Нормальный (доминантный) аллель этого гена будет обозначаться как А (он даёт функциональный продукт, который обуславливает разрушение хлорофилла и, соответственно, жёлтый цвет семян), а мутантный рецессивный аллель (ген не даёт функционального продукта, и семена остаются зелёными) как а. Именно поэтому такая мутация, когда теряется способность кодировать полноценный продукт, и обуславливает появление рецессивной формы этого гена.
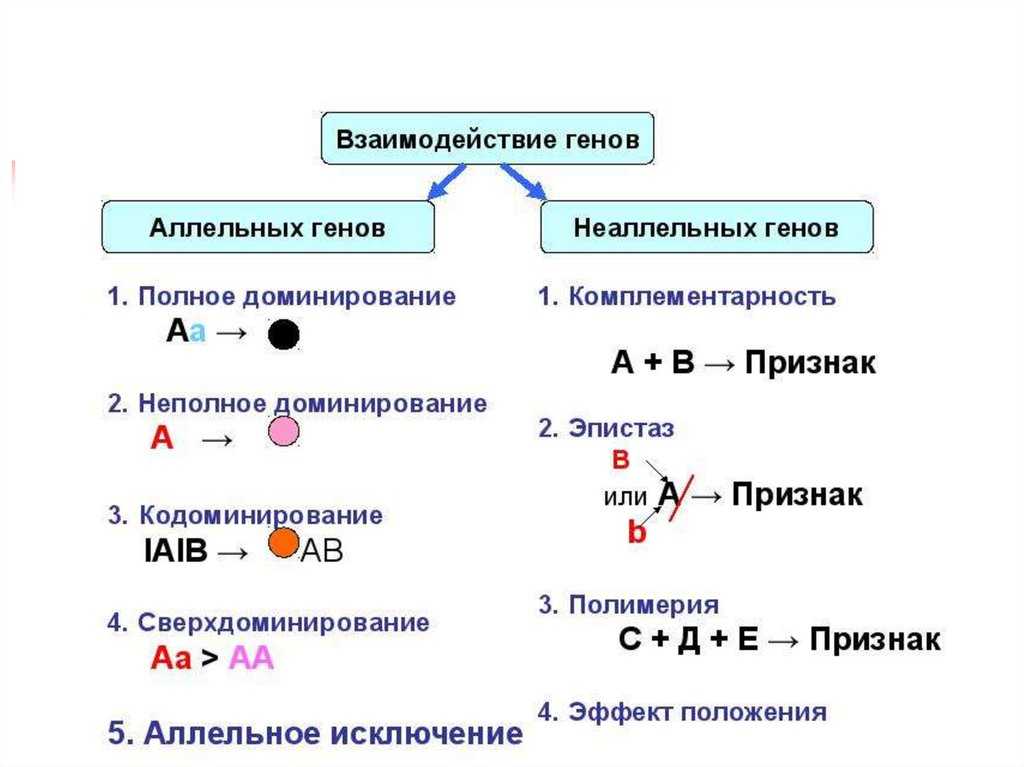
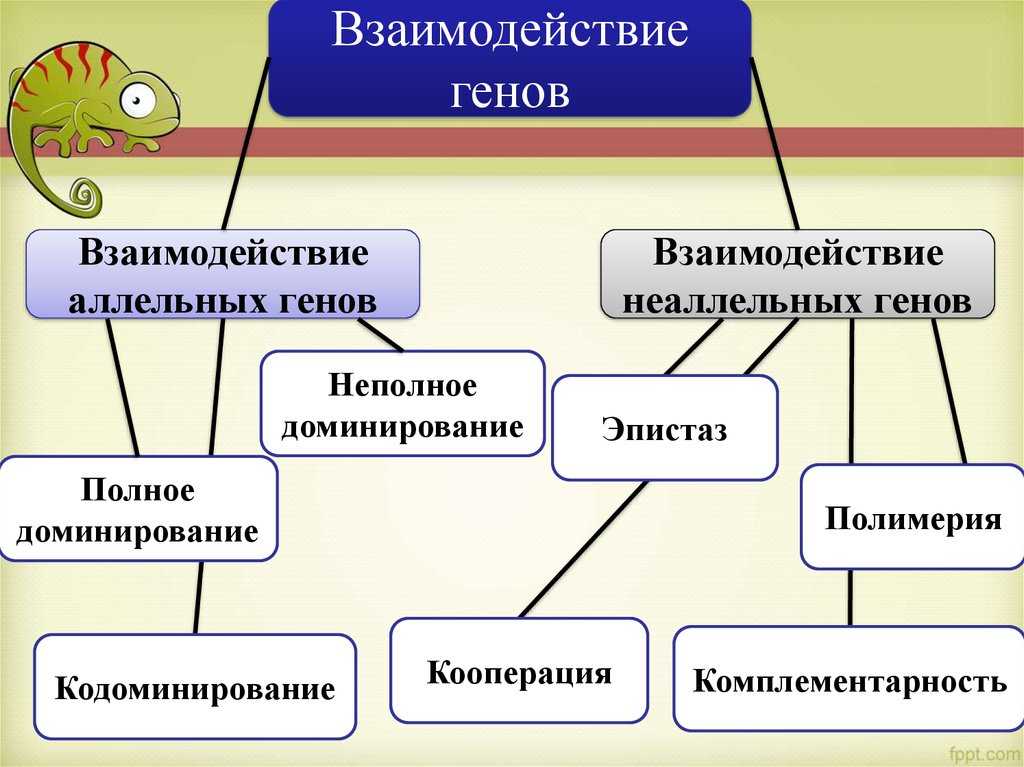
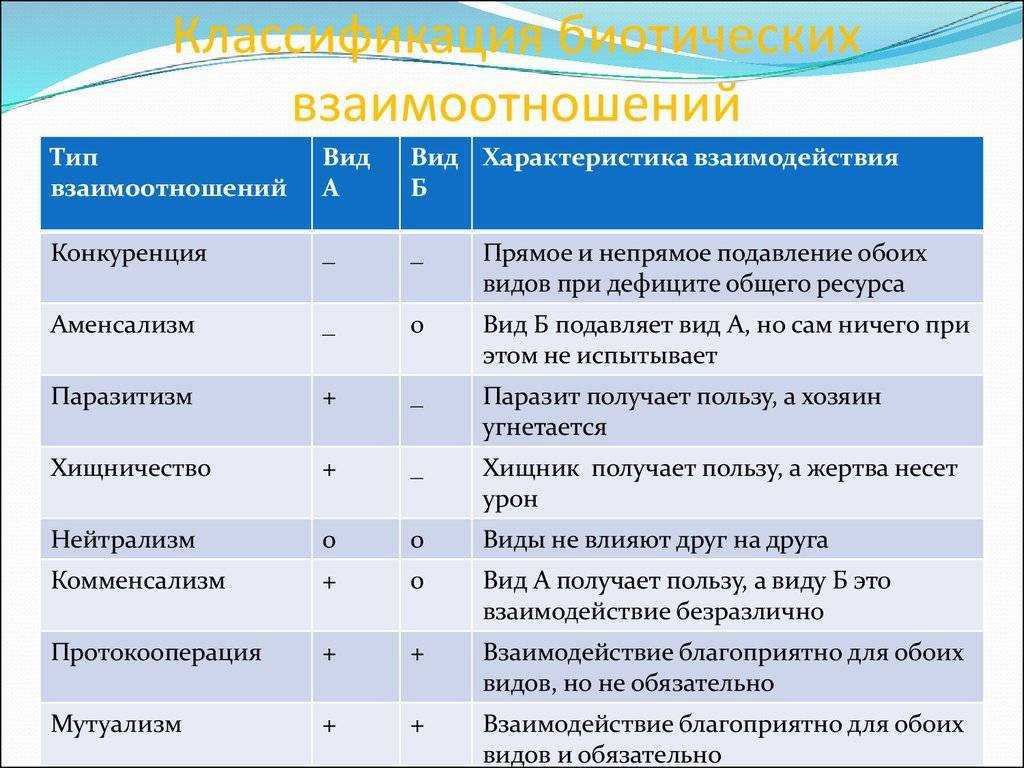
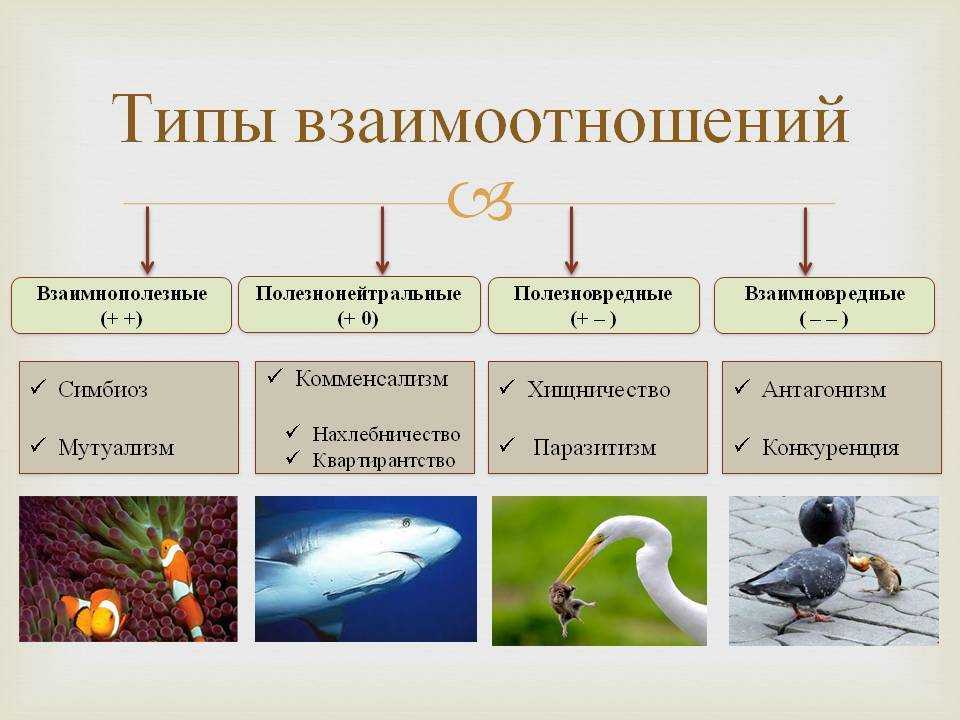
Типы взаимодействия аллельных генов
Различают три типа взаимодействия аллельных генов: полное доминирование, неполное доминирование, кодоминирование.
- Полное доминирование — явление, когда доминантный ген полностью подавляет работу рецессивного гена, в результате чего развивается доминантный признак.
- Неполное доминирование — явление, когда доминантный ген не полностью подавляет работу рецессивного гена, в результате чего развивается промежуточный признак.
- Кодоминирование (независимое проявление) — явление, когда в формировании признака у гетерозиготного организма участвуют оба аллеля.
У человека с серией множественных аллелей представлен ген, определяющий группу крови. При этом гены, обусловливающие группы крови A и B, являются кодоминантными по отношению друг к другу и оба доминантны по отношению к гену, определяющему группу крови 0.
![4.3. взаимодействие генов. генетика человека с основами общей генетики [учебное пособие]](https://lesniepolyani.ru/wp-content/uploads/3/a/7/3a7c7cd41af49781d9f9f16809225d58.jpeg)



3.2 Эпистаз
Вернемся к анализу взаимодействия генов рrи stу дрозофилы. Соотношение фенотипических классов в F2 можно представить себе и как следствие того, что
рецессивная аллель рrв
гомозиготе препятствует проявлению доминантной аллели st+. Точно так же рецессивная аллель stв гомозиготе препятствует проявлению
доминантной аллели рr+. Действительно, то, что известно о
генетическом контроле синтеза бурого глазного пигмента у дрозофилы, вполне
соответствует предложенному здесь объяснению. Такой тип взаимодействия носит
название эпистатического, или эпистаза, и условно изображается: рr> st+и st > pr+. В данном случае рецессивная аллель рrэпистатична по отношению к
доминантной аллели st+, a stэпистатична по отношению к рr+. Данный случай взаимодействия генов называют также двойным
рецессивным эпистазом.
По изменению числа и соотношения классов дигибридного
расщепления в F2 рассматривают несколько типов эпистатических
взаимодействий: простой рецессивный эпистаз (а > В; а> Ь или b> А; Ь > а), который
выражается в расщеплении 9:3:4; простой доминаданый эпистаз (А> В; А> bили В > А; B> а) с расщеплением 12:3:1 и т.д.
Один ген, подавляющий действие другого, называют эпистатическим
геном, ингибитором или супрессором. Подавляемый ген носит
название гипостатического.
Как уже показано, констатация того или иного типа
взаимодействия генов в дигибридном скрещивании условна. Тем не менее, при
кажущемся нарушении закона независимого наследования (появлении неожиданных
классов в расщеплении или уменьшении числа классов), связанного с
взаимодействием двух генов, всегда можно свести наблюдаемые соотношения в F2 к классическому 9:3:3:1
При этом
важно понять, какие классы объединились, и тогда интерпретировать тип
взаимодействия
Необходимо также отметить, что само словосочетание «взаимдействие
генов» условно. В действительности взаимодействуют продукты генов, а не сами
гены, так что правильнее было бы говорить о взаимодействии фенов, а не о
взаимодействии генов. Отсюда понятно, что судить о том, с каким скрещиванием
имеет дело экспериментатор: моногибридным, дигибридным или полигибридным —
можно только на основании результатов полного гибридологического анализа.
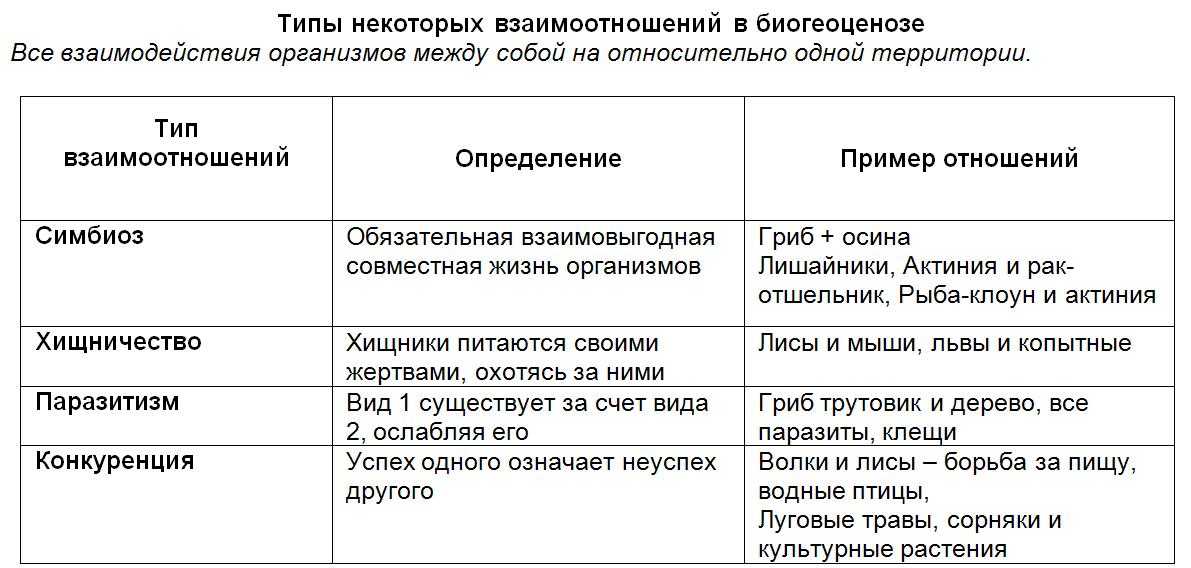
Типы взаимодействия неаллельных генов
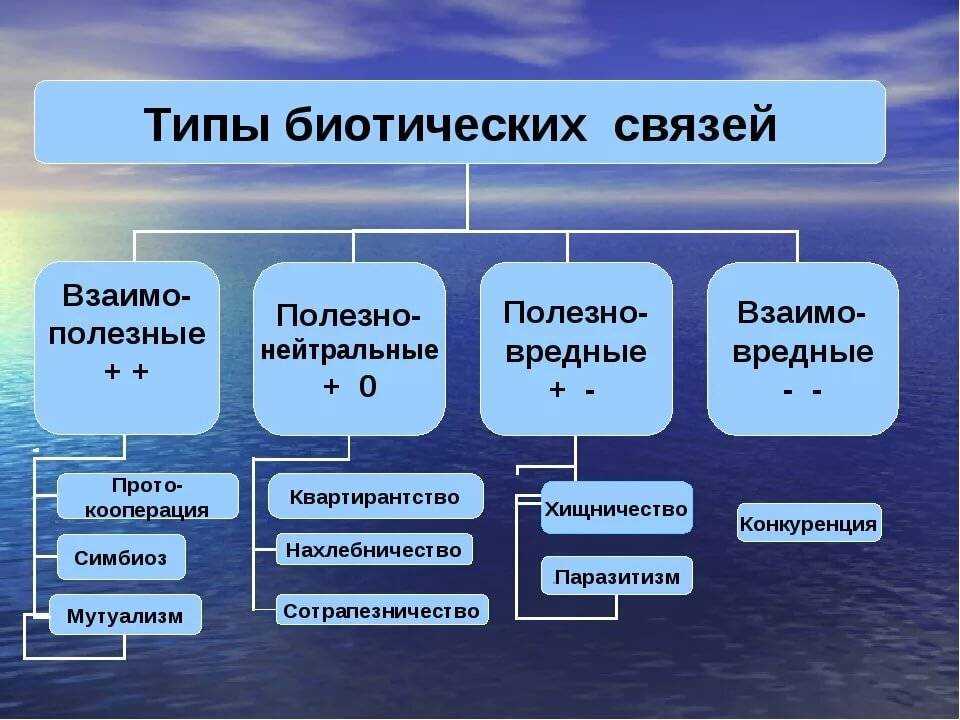
Различают четыре типа взаимодействия неалльлельных генов: кооперацию, комплементарность, эпистаз и полимерию.
Кооперация — явление, когда при взаимном действии двух доминантных неаллельных генов, каждый из которых имеет свое собственное фенотипическое проявление, происходит формирование нового признака.
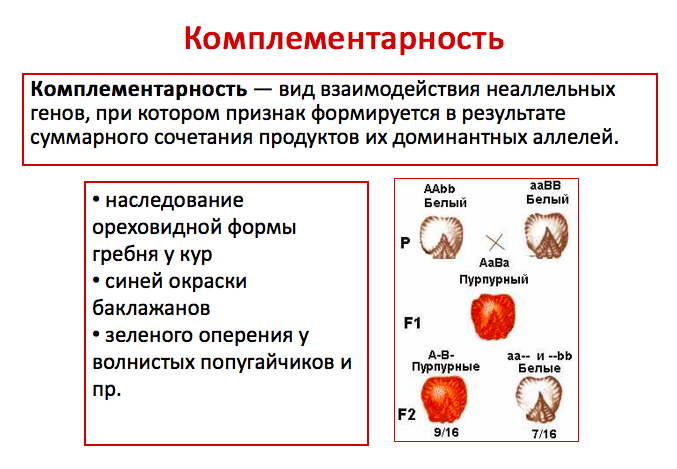
Комплементарность — явление, когда признак развивается только при взаимном действии двух доминантных неаллельных генов, каждый из которых в отдельности не вызывает развитие признака.
Эпистаз — явление, когда один ген (как доминантный, так и рецессивный) подавляет действие другого (неаллельного) гена (как доминантного, так и рецессивного).
Ген-подавитель (супрессор) может быть доминантным (доминантный эпистаз) или рецессивным (рецессивный эпистаз).
Полимерия — явление, когда несколько неаллельных доминантных генов отвечают за сходное воздействие на развитие одного и того же признака.
Чем больше таких генов присутствуют в генотипе, тем ярче проявляется признак. Явление полимерии наблюдается при наследовании количественных признаков (цвет кожи, масса тела, удой коров).
В противоположность полимерии наблюдается такое явление, как плейотропия — множественное действие гена, когда один ген отвечает за развитие нескольких признаков.
Генотип как целостная система. Взаимодействие генов
Вы познакомились с основными закономерностями наследования признаков у организмов.
Знания о цитологических основах наследственности развивались постепенно. Так, Г.Мендель, ничего не зная о генах и хромосомах, гениально сформулировал следующие положения:
- каждый признак организма контролируется парой наследственных задатков, или факторов (сейчас их называют генами);
- наследственные факторы (гены) могут существовать в двух состояниях: доминантном и рецессивном;
- гаметы генетически чисты, то есть содержат по одному наследственному фактору (гену) из каждой аллельной пары;
- при образовании зиготы наследственные факторы (гены) не смешиваются, остаются в «чистом» виде.
В 1909 г. Иогансен заменил термин «фактор» термином «ген».
В 1910-1920 гг. американский ученый Томас Морган сформулировал хромосомную теорию наследственности, согласно которой гены располагаются в хромосомах в линейном порядке, каждый ген занимает в хромосоме определенное место и влияет на формирование определенного признака.
Дальнейшие исследования внесли существенные поправки и дополнения в хромосомную теорию наследственности и углубили знания о гене.
Большинство генов влияет не на один признак организма, а на несколько, то есть обладает множественным действием.
Например, один из рецессивных генов у мухи дрозофилы обусловливает белую окраску глаз, низкую плодовитость и небольшую продолжительность жизни. У мышей под влиянием рецессивного гена в потомстве появляются карликовые особи, которые отличаются от нормальных мышей более тупыми мордочками, короткими усиками и хвостами, меньшей продолжительностью жизни.
В генотипе многих организмов имеются рецессивные летальные гены, одним из проявлений которых является гибель организма до полного завершения его развития, если летальный ген находится в гомозиготном состоянии.
Например, ген, обусловливающий отсутствие хлорофилла, приводит к гибели гомозиготных рецессивных проростков кукурузы.
В настоящее время установлено, что гены в генотипе тесно взаимосвязаны и взаимодействуют. Поэтому генотип рассматривают не как арифметическую сумму генов, а как целостную систему взаимодействующих генов. Полное и неполное доминирование обусловлено взаимодействием двух аллелей одного гена (А и а или B и b), которые определяют проявление у организма какого-то одного признака.
Это аллель 1000 ное взаимодействие генов.
Существуют и другие разнообразные взаимодействия генов. Так, гены-модификаторы ослабляют или усиливают действие других генов.
От их взаимодействия зависит, например, пятнистая окраска шерсти у животных: у одних особей пятнистость выражена в большей степени, у других в меньшей.
Некоторые неаллельные гены, одновременно находясь в генотипе, обусловливают развитие нового признака у организма. Например, при скрещивании черного и белого кроликов появляется потомство, которое имеет серую окраску шерсти.
При скрещивании гибридных особей между собой в потомстве происходит необычное расщепление по фенотипу: 9 серых : 3 черных : 4 белых.
Почему при скрещивании черного и белого кроликов в потомстве появляется новый признак — серая окраска шерсти?
Дело в том, что у кроликов за окраску шерсти отвечает аллельная пара генов Аа (А — черная, а — белая окраска шерсти), а за распределение пигмента по длине волоса — аллельная пара генов Вb (В — пигмент находится у корня волос, b — пигмент распределяется равномерно по длине волоса).
Теперь понятно, что черными будут кролики, у которых в генотипе ген А сочетается с геном b (Aabb, Aabb).
Кролики с генотипами ааВb, aaBB, aabb будут белыми, так как пигмент у них отсутствует. Серая окраска шерсти появляется только в том случае, если в генотипе соединились два доминантных неаллельных гена — А и В.

Это значит, что в клетках образуется пигмент (есть ген А), но он скапливается у основания волос (АаВb, ААВb, AaBB, AABB).
Большинство признаков организма формируется благодаря взаимодействию нескольких генов, которые отвечают за один и тот же признак, но не являются аллельными.
К таким признакам относят рост, массу тела животных, жирность и надои молока коров, яйценоскость кур. Эти гены принято обозначать одной и той же буквой со значками 1, 2, 3 и т. д. Цвет кожи у человека, например, обусловлен двумя неаллельными генами A1 и А2. Он зависит от числа доминантных генов в генотипе: чем их больше, тем темнее кожа. Люди, генотип которых А1A1А2А2, будут иметь темную кожу, а кожа людей с генотипом а1а1а2а2 светлая.
Люди с генотипами А1a1А2a2, А1a1a2a2 имеют различные оттенки кожи — от более темной до почти светлой.
Полезно знать
VetConsult+, 2015. Все права защищены. Использование любых материалов, размещённых на сайте, разрешается при условии ссылки на ресурс. При копировании либо частичном использовании материалов со страниц сайта обязательно размещать прямую открытую для поисковых систем гиперссылку, расположенную в подзаголовке или в первом абзаце статьи.
Из данного видеоурока вы узнаете о том, что взаимодействие между генами происходит путём взаимодействия продуктов, которые эти гены кодируют. Познакомитесь с видами взаимодействия генов в генотипе, взаимодействием аллельных генов и неаллельных генов а также с множественным действием одного гена. В данном уроке приводятся следующие понятия: взаимодействие аллельных генов, полное доминирование, неполное доминирование, кодоминирование, аллельное исключение, взаимодействие неаллельных генов, комплементарность, эпистаз, полимерия, плейтропность
Частота аллелей
Частота встречаемости аллельных генов в популяции может быть различной. Обычно рецессивные гены редки и по-сути являются мутациями доминантного аллеля. Многие мутации являются вредными. Однако именно мутантные гены создают материал для действия естественного отбора и как следствие процесса эволюции.
В гипотетической идеальной популяции (в которой не действует естественный отбор, которая имеет неограниченно большой размер, изолирована от других популяций и так далее) частота генотипов (по тому или иному гену) не меняется и подчиняется закону Харди-Вайнберга. Согласно этому закону распределение генотипов в популяции будет укладываться в уравнение: p2 + 2pq + q2 = 1. Здесь p и q — частоты (выраженные в долях единицы) аллелей в популяции, p2 и q2 — частоты соответствующих гомозигот, а 2pq — частота гетерозигот.
Причины отклонений от менделевских расщеплений
Менделевские расщепления в принципе могут быть реализованы только для видов живых организмов с нормальным половым процессом, диплоидностью обоих полов и при условии отсутствия нарушений в мейозе. Но и при выполнении указанных условий случаи отклонения от менделевских расщеплений достаточно многочисленны. При этом каких-либо исключений нет, а условия менделирования не нарушаются — просто на них накладываются другие эффекты.

Три таких эффекта — взаимодействие неаллельных генов (комплементация, кумулятивная и некумулятивная полимерия), сцепление групп генов в одной хромосоме и расположение гена в половых хромосомах. Среди других причин выделяются такие:
- Летальность некоторых комбинаций аллелей. Например, при скрещивании жёлтых (гетерозиготных) мышей, расщепление на жёлтые и чёрные (рецессивные гомозиготы) всегда составляет 2:1. Подобный феномен наблюдается также при скрещивании платиновых лисиц. Если проверить беременных самок, то оказывается, что четверть эмбрионов погибает ещё до рождения: погибают гомозиготы по доминантному аллелю, поэтому и наблюдается отклонение от ожидаемого расщепления 3:1 (отсутствует один из четырёх генотипических классов). Таким образом, в этой задаче доминантный аллель является на самом деле доминантным только по окрасу шерсти. Что касается жизнеспособности, этот самый аллель является рецессивным. Ситуация, когда один ген влияет на несколько признаков одновременно (окрас шерсти и жизнеспособность), является одним из примеров плейотропии.
- Существуют аллели, которые фенотипически проявляются только у определённой группы организмов, содержащих их в генотипе. Для характеристики такого вариабельного проявления генов используют понятие пенетрантности и экспрессивности. Согласно определению, мерой пенетрантности является группа особей, характеризующихся определённым фенотипом, среди всех особей с одинаковым генотипом. Некоторые наследственные болезни человека, например, подагра, характеризуются неполной пенетрантностью, то есть не все особи, которые несут мутацию, болеют. Экспрессивность отображает силу проявления мутантного гена в фенотипе. Так, дрозофилы, гомозиготные по мутантному аллелю eyeless, могут иметь разное количество фасеток глаз — от нормального количества до полного их отсутствия. Способность генотипа проявляться по-разному в зависимости от внешних условий отражает норму реакции — определённый диапазон проявления (изменчивость), в границах которого признаки могут модифицироваться в ответ на вариации условий развития организма.
- Некоторые признаки присущи особям в течение не всей жизни, а возникают только в определённый период: характер расщепления может иметь взаимосвязь с возрастом. Например, у человека имеются наследственные синдромы, которые проявляются только в преклонном возрасте (синдром Альцгеймера, болезнь Хантингтона).
- К отклонениям от ожидаемых менделевских расщеплений приводит также явление импринтинга. Основой его является так называемая эпигенетическая наследственность — может наследоваться от родителей не просто ДНК, а хроматина, который несёт определённые химические маркеры. От них зависит структурное состояние участка и, соответственно, активность некоторых генов. Вследствие такого эффекта проявление определённого гена может быть обусловлено тем, от кого из родителей хромосома была получена.
- В природе существует много видов полового диморфизма, и пол играет большую роль в проявлении ряда признаков.
Молекулярные механизмы взаимодействий генов
Взаимодействие аллельных генов:
* Полное доминирование: Явление, при котором доминантный ген полностью подавляет рецессивный. Гетерозиготные организмы по фенотипу всегда точно соответствуют родителю, гомозиготному по доминантному гену.
* Неполное доминирование: Явление, при котором доминантный ген не полностью подавляет работу рецессивного, в результате развивается промежуточный признак.
Примером может служить окраска цветка у растения ночная красавица.
У которого при скрещивании красных и белых цветков среди гибридов первого поколения все цветки розовые.
* Множественный аллелизм: Явление, при котором ген представлен не двумя аллелями (как в случаях полного или неполного доминирования), а гораздо большим их числом.
Пример – трёхчленная серия аллелей, определяющая окраску меха у кошек. Чёрная окраска меха B доминирует над шоколадной b, которая, в свою очередь, доминирует над светло-коричневой b1. (B>b>b1).
* Кодоминирование: Явление независимого друг от друга проявления обоих аллелей в фенотипе гетерозиготы, иными словами – отсуствие доминантно-рецессивных отношений между аллелями.
Пример – взаимодействие аллелей (IA и IB), определяющих четвёртую группу крови человека.
* Сверхдоминирование: более сильное проявление признака в гетерозиготе, а не в гомозиготе.
Так, у дрозофилы имеется рецессивный летальный ген, гетерозиготы по которому обладают большей жизнеспособностью, чем доминантные гомозиготы.
Взаимодействие неаллельных генов:
* Комплиментарность (лат. – «комплиментум» — дополнение) – один из видов взаимодействия неаллельных генов, при котором эти гены дополняют действие друг друга, и признак формируется при одновременном действии обоих генов. У душистого горошка есть доминантный ген A, обуславливающий синтез пропигмента – бесцветного предшественника пигмента. (Его рецессив «а» определяет отсутствие пропигмента).
Другой доминантный ген, B, определяет синтез фермента, под действие которого из пропигмента образуется пигмент. (Его рецессив «b» определяет отсутствие фермента). Теперь рассмотрим эксперимент по скрещиванию двух растений с генотипами AAbb и aaBB. В обоих случаях растения имеют белые цветки. В F1 образуются дигетерозиготы, у которых есть и пропигмент, и фермент.
Поэтому в F1 все растения будут иметь розовые цветки.
* Эпистаз— тип взаимодействия генов, при котором один ген подавляет действие другого. Пример: У тыквы доминантный ген Y вызывает появление жёлтой окраски плодов, а её рецессив y – зелёной. Кроме того, имеется доминантный ген W, подавляющий действие любой окраски, следовательно с его присуствием плоды будут белые, в то время как его рецессив w не препятствует проявлению окраски.
Так, например, у растений с генотипами YyWw, yyWw плоды будут белыми, а у растений с генотипами yyww YYww зелёными и жёлтыми соответственно.
* Полимерия— тип взаимодействия проявление генов, при котором проявление конкретного признака зависит от количества доминантных генов, вносящих вклад в его развитие.
Например, при скрещивании краснозерных пшениц с белозерными было установлено, что растения с генотипом А1А1А2А2 имеют красные зерна, растения а1а1а2а2 – белые зерна, растения с тремя доминантными генами – красноватую окраску, а растения с двумя и одним геном – более бледную окраску.
3.3 Полимерия
Наряду с комплементарным и эпистатическим принято также
рассматривать взаимодействие генов по типу полимерии. В этом случае разные гены
как бы дублируют действие друг друга, и одной доминантной аллели любого из
взаимодействующих генов достаточно для проявления изучаемой фенотипической
характеристики. Так, при скрещивании растений пастушьей сумки с треугольными плодами
(стручками) и с овальными плодами в F1образуются растения с плодами треугольной формы. При
их самоопылении в F2 наблюдается расщепление на растения
с треугольными и овальными стручками в соотношении 15:1. Это объясняется тем,
что существуют два гена, действующих эднозначно. В этих случаях их обозначают
одинаково (А1 и А2). Тогда все генотипы: А1
— А2 -, А1 — а2а2,
а2а2А2— будут иметь
одинаковую фенотипическую характеристику — треугольные стручки, и только
растения а1а1а2а2 будут отличаться — образовывать
овальные стручки. Это случай так называемый некумулятивной полимерии.


Однозначные, или полимерные, гены могут действовать и по типу
кумулятивной полимерии. Так, шведский генетик Г. Нильсон-Эле в 1908 г.
описал серию однозначно действующих генов, которые определяют окраску
эндосперма зерен пшеницы. При этом интенсивность окраски зерен оказалась
пропорциональной числу доминантных аллелей разных генов в тригибридном
скрещивании. Наиболее окрашенными были зерна А1А1А2А2А3А3,
а зерна а1а1а2а2а3а3 не
имели пигмента. Между этими крайними типами при расщеплении в F2 наблюдались промежуточные варианты в соотношении
1:6:15:20:15:6:1.
По типу кумулятивной полимерии наследуются многие количественные
признаки, например цвет кожи у человека; молочность, яйценоскость, масса и
другие признаки сельскохозяйственных животных; длина колоса у злаков,
содержание сахара в корнеплодах сахарной свеклы и др. Изучением наследования
таких признаков занимается специальный раздел генетики — генетика
количественных признаков, которая важна прежде всего для селекции и разработки
проблем микроэволюции.
Экспрессия генов: что это?
Экспрессия генов — это процесс, в ходе которого наследственная информация от гена (последовательности нуклеотидов ДНК) преобразуется в функциональный продукт — РНК или белок.
Экспрессия генов может регулироваться на всех стадиях процесса: и во время транскрипции, и во время трансляции, и на стадии посттрансляционных модификаций белков.
Регуляция экспрессии генов позволяет клеткам контролировать собственную структуру и функцию и является основой дифференцировки клеток, морфогенеза и адаптации.
Экспрессия генов является субстратом для эволюционных изменений, так как контроль за временем, местом и количественными характеристиками экспрессии одного гена может иметь влияние на функции других генов в целом организме.
Экспрессивность и пенетрантность
Под экспрессивностью понимают степень выраженности определенного признака.
Внешняя среда и гены-модификаторы могут изменить экспрессию гена, т. е. выражение признака. Изменчивость проявления мутантного гена у разных особей — довольно частое явление. Например, у потомства дрозофилы — мутантных «безглазых» мух с сильно редуцированным количеством фасеток — содержание их варьирует от почти полного отсутствия до половины нормы. Пенетрантность гена — это доля особей, у которых проявляется ожидаемый фенотип.
При полной пенетрантности (100 %) мутантный ген проявляет свое действие у каждой особи. При неполной пенетрантности (меньше 100 %) ген проявляется фенотипически не у всех особей. Экспрессивность и пенетрантность гена в значительной степени зависят, по-видимому, от влияния генов-модификаторов и условий развития особей.
Норма реакции — способность генотипа формировать в онтогенезе, в зависимости от условий среды, разные фенотипы.
Она характеризует долю участия среды в реализации признака и определяет модификационную изменчивость вида.
Чем шире норма реакции, тем больше влияние среды и тем меньше влияние генотипа в онтогенезе. Один и тот же ген в разных условиях среды может реализоваться в несколько проявлений признака (фенов). В каждом конкретном онтогенезе из спектра проявлений признака реализуется только один.
Аналогично один и тот же генотип в разных условиях среды может реализоваться в целый спектр потенциально возможных фенотипов, но в каждом конкретном онтогенезе реализуется только один фенотип.
Под наследственной нормой реакции понимают максимально возможную ширину этого спектра: чем он шире, тем шире норма реакции. Фенотипическое значение любого количественного признака (Ф) определяется, с одной стороны, его генотипическим значением (Г), с другой стороны — влиянием среды (С): Ф = Г + С.