

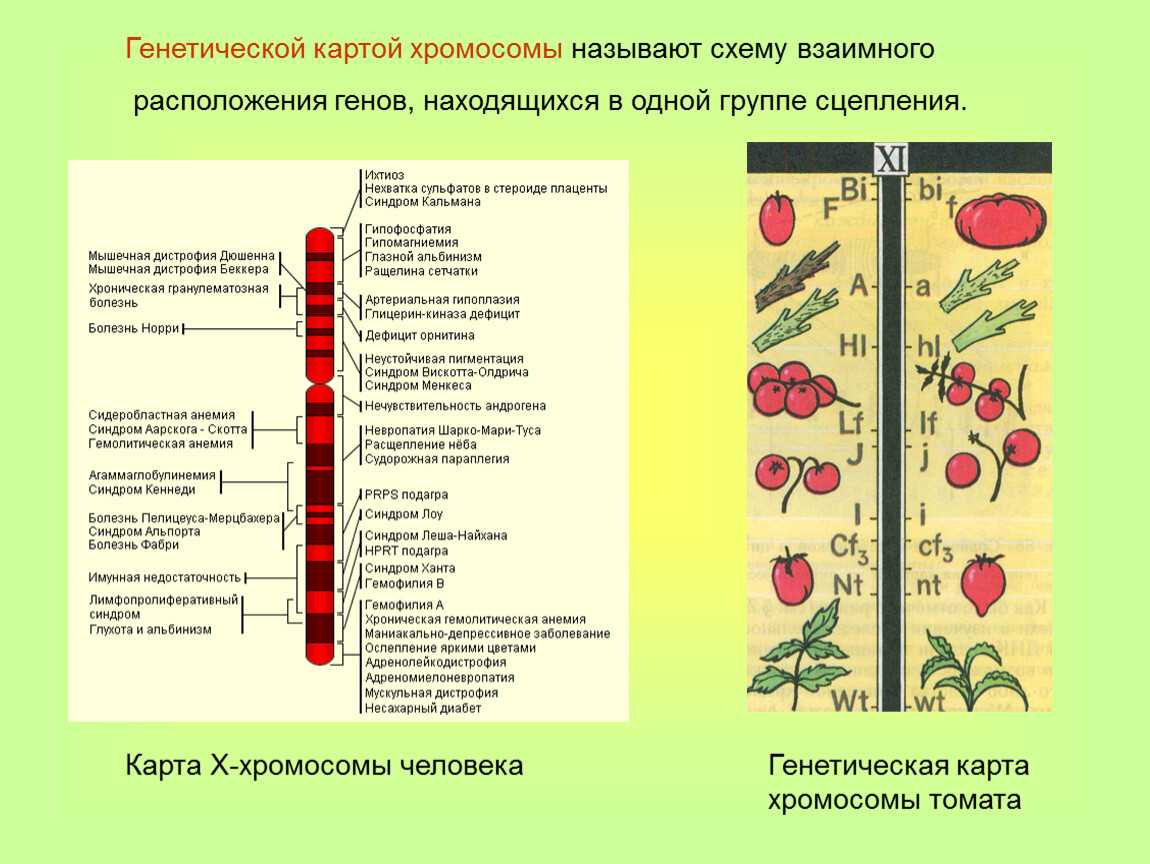
Генетические карты хромосом
Большая советская энциклопедия. — М.: Советская энциклопедия . 1969—1978 .
Смотреть что такое «Генетические карты хромосом» в других словарях:
ГЕНЕТИЧЕСКИЕ КАРТЫ ХРОМОСОМ — схемы относительного расположения генов в хромосомах, позволяющие предсказывать характер наследования изучаемых признаков организмов … Большой Энциклопедический словарь
генетические карты хромосом — схемы относительного расположения генов в хромосомах, позволяющие предсказывать характер наследования изучаемых признаков организмов. * * * ГЕНЕТИЧЕСКИЕ КАРТЫ ХРОМОСОМ ГЕНЕТИЧЕСКИЕ КАРТЫ ХРОМОСОМ, схемы относительного расположения генов в… … Энциклопедический словарь
ГЕНЕТИЧЕСКИЕ КАРТЫ ХРОМОСОМ — схемы относительного расположения генов в хромосомах, позволяющие предсказывать характер наследования изучаемых признаков организмов … Естествознание. Энциклопедический словарь
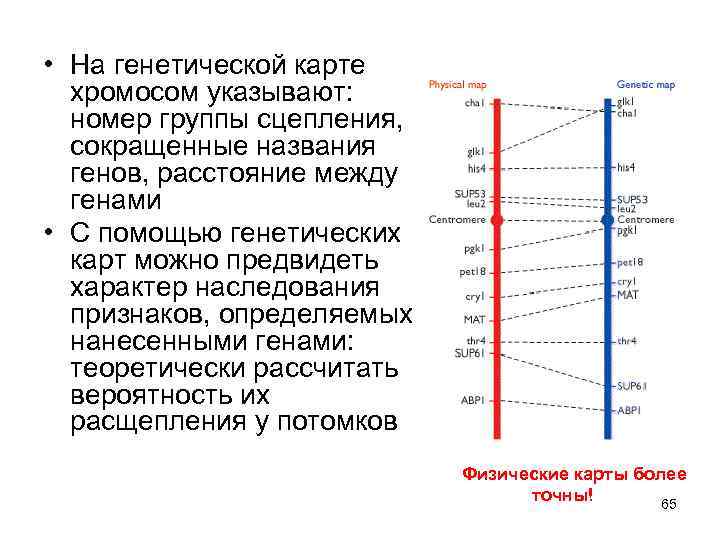
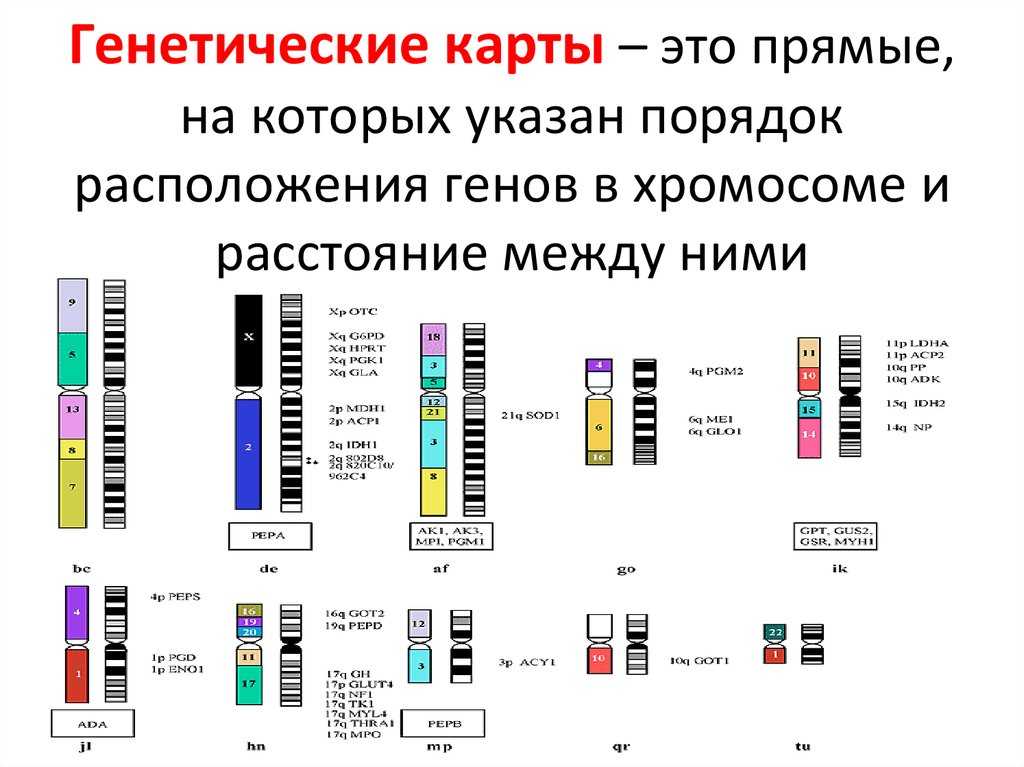
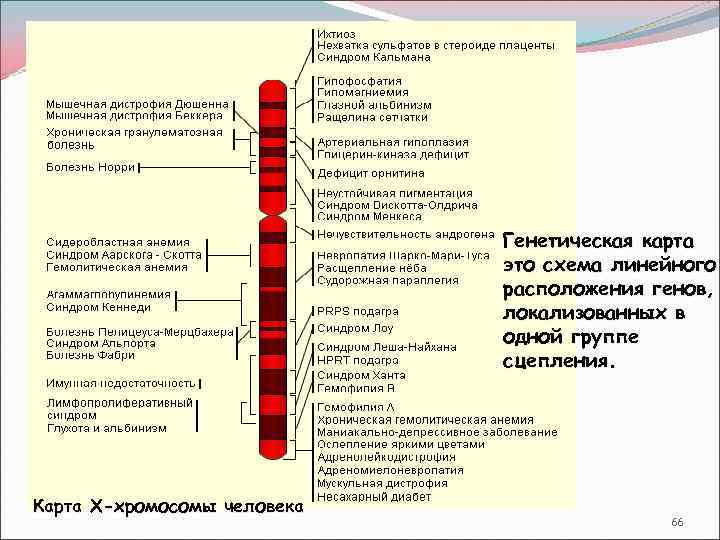
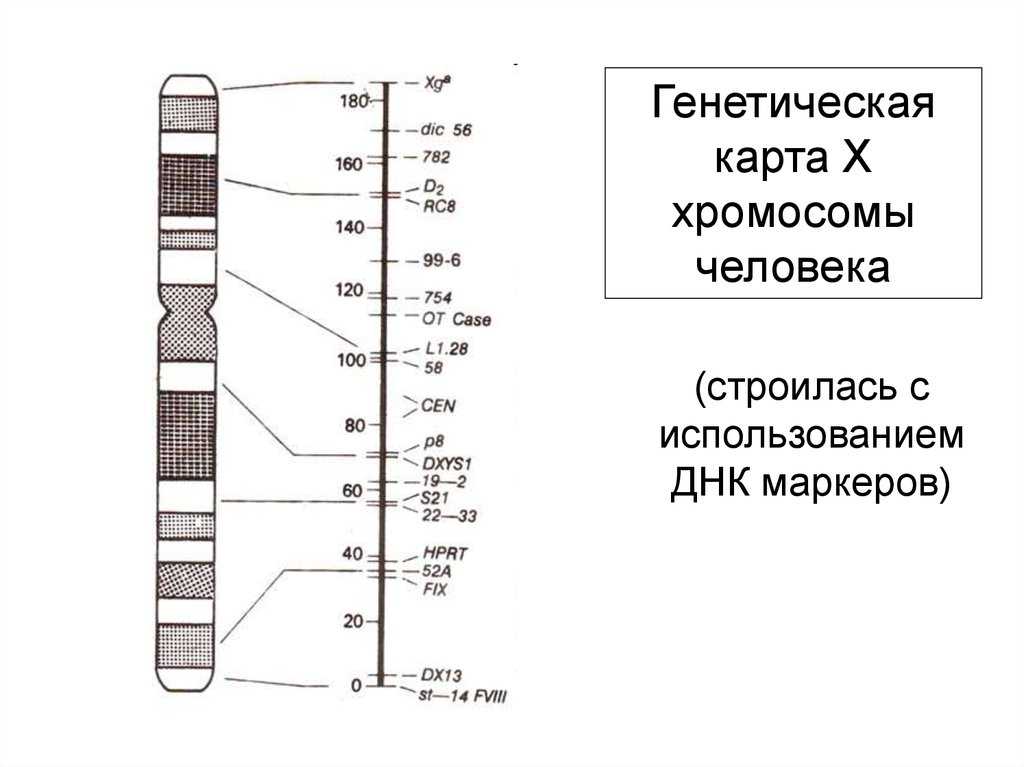
Карты хромосом — Графическое представление нормального человеческого кариотипа. Генетические карты хромосом это схема взаимного расположения и относительных расстояний между генами определенных хромосом, находящихся в од … Википедия
Цитологические карты хромосом — схематическое изображение хромосом (См. Хромосомы) с указанием мест фактического размещения отдельных генов, полученное с помощью цитологических методов. Ц. к. х. составляют для организмов, для которых обычно уже имеются Генетические… … Большая советская энциклопедия
Хромосомные карты генетические к — Хромосомные карты, генетические к. * храмасомныя карты, генетычныя к. * chromosome maps or genetic m. графическое изображение хромосом, как правило, в виде прямых линий, на которых указаны гены/локусы определенной группы сцепления (см.) и… … Генетика. Энциклопедический словарь
Генетика — I Генетика (от греч. génesis происхождение) наука о законах наследственности и изменчивости организмов. Важнейшая задача Г. разработка методов управления Наследственностью и наследственной Изменчивостью для получения нужных человеку форм… … Большая советская энциклопедия
Генетика — I Генетика (от греч. génesis происхождение) наука о законах наследственности и изменчивости организмов. Важнейшая задача Г. разработка методов управления Наследственностью и наследственной Изменчивостью для получения нужных человеку форм… … Большая советская энциклопедия
генетика — , и; ж. . Наука о законах наследственности и изменчивости организмов. Г. человека. Г. растений. Медицинская г. Космическая г. * * * генетика (от греч. génesis происхождение), наука о… … Энциклопедический словарь
Хромосомная теория наследственности — Хромосомная теория наследственности теория, согласно которой хромосомы, заключённые в ядре клетки, являются носителями генов и представляют собой материальную основу наследственности, то есть преемственность свойств организмов в ряду… … Википедия
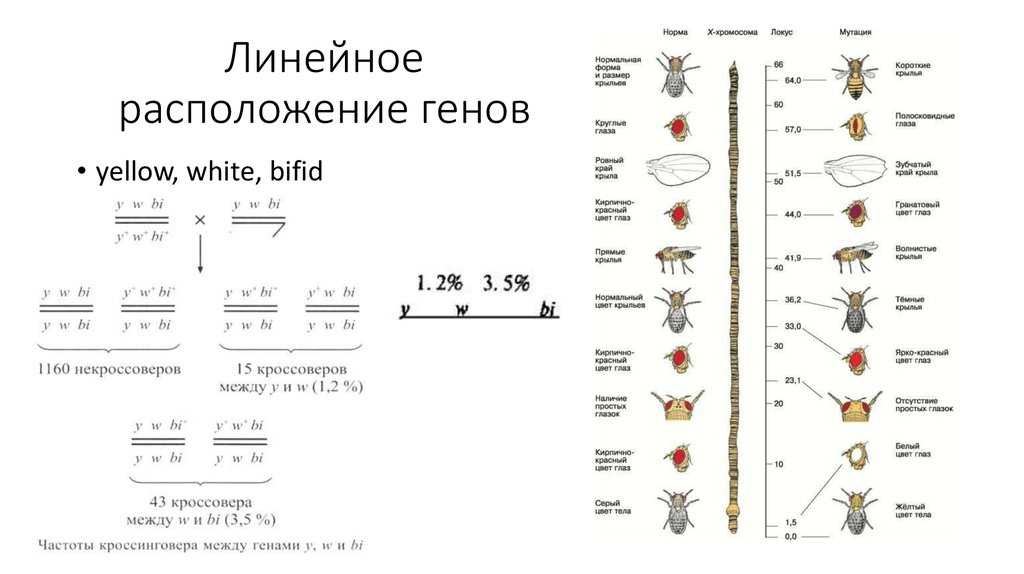
Вопрос 64. Линейное расположение генов. Генетические карты
1.
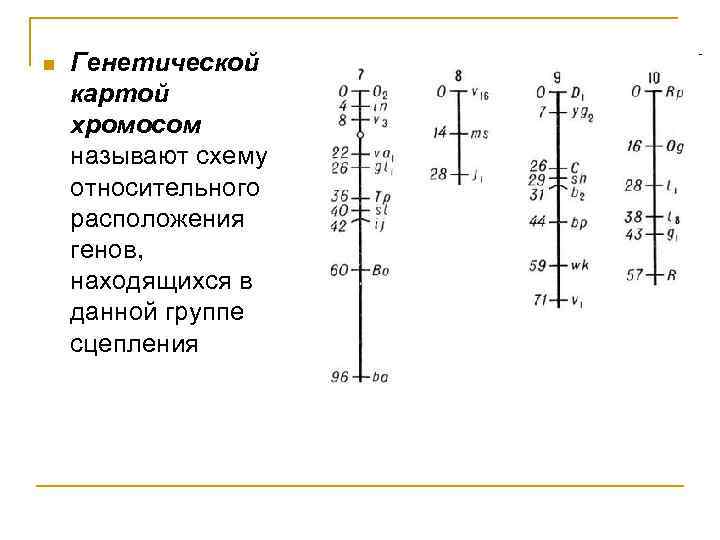
Основа принципа построения генетических карт
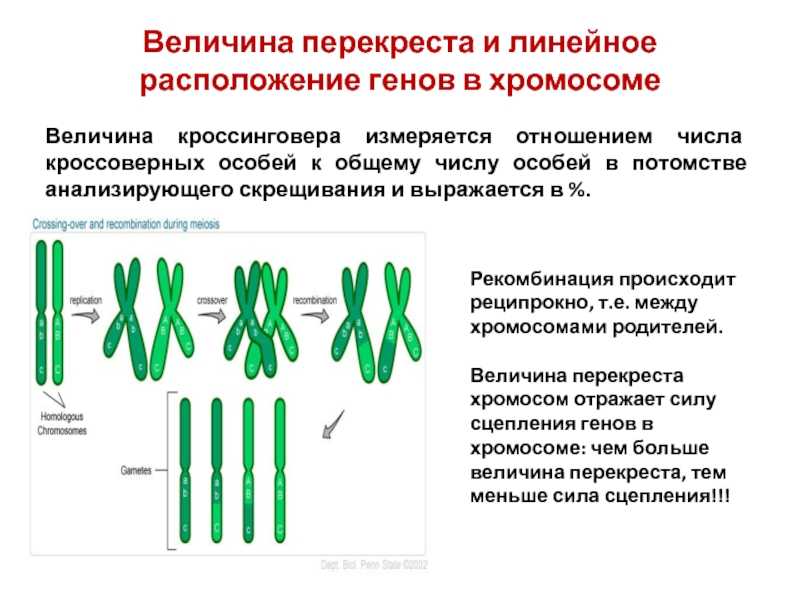
Существование кроссинговера побудило Моргана разработать в 1911–1914 гг. принцип построения генетических карт хромосом. В основу этого принципа положено представление о расположении генов по длине хромосомы в линейном порядке. За единицу расстояния между двумя генами условились принимать 1 % перекреста между ними.
Допустим, что к одной группе сцепления относятся гены А и В. Между ними обнаружен перекрест в 10 5. Следовательно, гены А и В находятся на расстоянии 10 единиц. Допустим далее, что к этой же группе сцепления относится ген С. Чтобы узнать его место в хромосоме, необходимо выяснить, какой процент перекреста он дает с обоими из двух уже известных генов. Например, если с А он дает 3 % перекреста, то можно предположить, что ген С находится либо между А и В, либо с противоположной стороны, то есть А расположен между В и С. Если между В и С кажется перекрест 7 %, то на хромосоме их следует расположить в таком порядке, как на верхней схеме. Если между В и С перекрест составит 13 %, то расположение генов будет как на нижней схеме.
2.
В общей форме эту закономерность можно выразить следующей формулой : если гены А, В и С относятся к одной группе сцепления и расстояние между генами А и В равно нескольким единицам, а расстояние между В и С – одной единице, то расстояние между А и С может быть либо k +1, либо k-1.
Генетические карты хромосомы строятся на основе гибридологического анал иза . Однако найден способ построения и цитологических карт хромосом для дрозофилы. Дело в том, что в клетках личинок мух обнаружены гигантские хромосомы, превышающие размер хромосом из других клеток в 100–200 раз и содержащие в 1000 раз больше хромосом.
Оказалось, что в тех случаях, когда гибридологическим методом обнаруживались какие-либо нарушения наследования , соответствующие им изменения имели место и в гигантских хромосомах. Так, в результате сопоставления генетических и цитологических данных стало возможным построить цитологические карты хромосом. Это открытие подтверждает правильность тех принципов, которые были положены в основу построения генетических карт хромосом.
3.
Хромосомная теория наследственности
Основные положения хромосомной теории наследственности:
• I. Гены находятся в хромосомах. Каждая хромосома представляет собой группу сцепления генов. Число групп сцепления у каждого вида равно гаплоидному набору хромосом.
• II. Каждый ген в хромосоме занимает отдельное место (локус). Гены в хромосомах расположены линейно.






![Что такое цитологические карты хромосом и как они составляются? [1989 морозов е.и., тарасевич е.и., анохина в.с. - генетика в вопросах и ответах]](https://lesniepolyani.ru/wp-content/uploads/e/f/6/ef6a9bdb2a4f5748d25a6624651eac55.jpeg)
• III. Между гомологичными хромосомами происходит обмен аллельными генами.
• IV. Расстояние между генами в хромосоме пропорционально проценту кроссинговера между ними.
Генетика пола[]
Пол, как и любой другой признак организма, наследственно детерминирован. Важнейшая роль в генетической детерминации пола и в поддержании закономерного соотношения полов принадлежит хромосомному аппарату.
У раздельнополых организмов (животных и двудомных растении) соотношение полов обычно составляет 1:1, то есть мужские и женские особи встречаются одинаково часто. Это соотношение совпадает с расщеплением в анализирующем скрещивании, когда одна из скрещиваемых форм является гетерозиготной (Аа), а другая — гомозиготной по рецессивным аллелям (аа). В потомстве в этом случае наблюдается расщепление в отношении 1Аа:1аа. Если пол наследуется по такому же принципу, то вполне логично было бы предположить, что один пол должен быть гомозиготным, а другой — гетерозиготным. Тогда расщепление по полу должно быть в каждом поколении равным 1:1, что и наблюдается в действительности.
При изучении хромосомных наборов самцов и самок ряда животных между ними были обнаружены некоторые различия. Как у мужских, так и у женских особей во всех клетках имеются пары одинаковых (гомологичных) хромосом, но по одной паре хромосом они различаются. Так, у самки дрозофилы имеются две палочковидные хромосомы, а у самца — одна такая же палочковидная, а вторая, парная первой, — изогнутая. Такие хромосомы, по которым самцы и самки отличаются друг от друга, называют половыми хромосомами. Те из них, которые являются парными у одного из полов, называют X-хромосомами (например, у дрозофилы и млекопитающих) или Z-хромосомами (например, у птиц). Непарная половая хромосома, имеющаяся у особей только одного пола, была названа У-хромосомой (у дрозофилы и млекопитающих) или W-хромосомой (у птиц). Хромосомы, в отношении которых между самцами и самками нет различий, называют аутосомами. Следовательно, у дрозофилы особи обоих полов имеют по шесть одинаковых аутосом плюс две половые хромосомы (ХХ у самок и XY у самцов).
Пол, имеющий различные половые хромосомы (X и У), образует гаметы двух типов (половина с X-хромосомой и половина с У-хромосомой), то есть, является гетерогаметным, а пол, содержащий в каждой клетке одинаковые половые хромосомы (X-хромосомы), — гомогаметным.
Открытие половых хромосом и установление их роли в определении пола послужило важным доводом в пользу того, что хромосомы определяют признаки организма.
Слайд 32Хромосомы с множественными инверсиями используют при создании балансеров, т. е.
линий, позволяющих поддерживать летальные мутации и мутации по плодовитости. Один
из примеров — линия СLВ. Более надежными балансерами, т. е. содержащими несколько инверсий, являются линии Base, Binsn. Конструирование балансерных хромосом по существу представляет собой первый пример генетической инженерии.Другой пример балансеров — линия Су (загнутые крылья, летальность), в которой доминантная мутация сопряжена с длинной инверсией, захватывающей почти всю вторую хромосому. В потомстве от скрещивания гетерозигот по Су выживают только мухи родительских классов, т. е. линия сбалансирована, и исследуемая леталь /, постоянно в ней поддерживается в гетерозиготном состоянии.

Слайд 20Метод цитологических карт основан на использовании хромосомных перестроек. При облучении
и действии мутагенов в хромосомах часто наблюдаются потери (делеции) или
вставки (дупликации) небольших фрагментов, сравнимых по величине с одним или несколькими локусами.Например, можно использовать гетерозиготы по хромосомам, одна из которых будет нести группу следующих друг за другом доминантных аллелей, а гомологичная ей — группу рецессивных аллелей тех же генов ABCDE/abcde. Если в хромосоме с доминантными генами произошла утрата отдельных генов, например DE, то у гетерозиготы ABC/abcde будут проявляться рецессивные признаки de. На этом принципе основан метод перекрывающихся делеции, используемый при построении цитологических карт!!!

Вопрос 65. Основы молекулярной генетики. Обнаружение химической природы гена
1
. Носитель наследственной информации
Хромосомная теория наследственности закрепила за генами роль элементарных наследственных единиц, локализованных в хромосомах. Однако химическая природа гена долго еще оставалась неясной. В настоящее время известно, что носителем наследственной информации является ДНК. Убедительным доказательством того, что именно с ДНК связана передача наследственной информации, получена при изучении вирусов. Проникая в клетку, они вводят в нее лишь нуклеиновую кислоту с очень небольшой примесью белка, а вся белковая оболочка остается вне зараженной клетки. Следовательно, введенная в клетку ДНК передает генетическую информацию, необходимую для образования нового поколения вируса такого же типа.
2.
Роль нуклеиновой кислоты
Далее было обнаружено, что чистая нуклеиновая кислота вируса табачной мозаики может заразить растение, вызывая типичную картину заболевания. Более того, удалось искусственно создать вегетативные «гибриды» из вирусов, в которых белковый футляр принадлежал одному виду, а нуклеиновая кислота – другому. В таких случаях генетическая информация «гибридов» всегда в точности соответствовала тому виду вирусов, чья нуклеиновая кислота входила в состав «гибрида». Важным доказательством роли ДНК в передаче наследственной информации были получены на микробных клетках в явлениях трансформации и трансдукции.
3.
Трансформация – включение чужеродной ДНК в бактериальную клетку. Это перенос наследственной информации от одной клетки прокариот посредством ДНК бактерии-донора или клетки-донора. Трансформация происходит при обработке бактерии мертвыми клетками или экстрактами других штаммов. При этом бактерии приобретают определенные свойства и сохраняют их. Например, при обработке невирулентного (т. е. не вызывающего заболевания) штамма пневмококков экстрактом ДНК из вирулентного штамма он приобретал способность вызывать воспаление легких.
4.
Трансдукция заключается в том, что вирусы, покидая бактериальные клетки, в которых они паразитировали, могут захватывать с собой часть их ДНК и, перемещаясь в новые клетки, передавать новым хозяевам свойства прежних. Так, при инфицировании культуры неподвижных бактерий вирусами, размножающимися в подвижной культуре, некоторые из бактерий становятся подвижными. Следовательно, изменение наследственности бактерий при трансдукции достигается благодаря переносу ДНК от одних бактериальных клеток к другим с помощью ДНК или РНК вируса.
5.
Перенос генетического материала
Описан перенос генетического материала (ДНК) посредством вирусов (то есть трансдукции) у насекомых. Непигментированных личинок тутового шелкопряда заражали вирусами, которые до этого паразитировали в окружающих насекомых. В результате у части потомства, полученного от непигментированных, но зараженных особей, появилась окраска. И в этом случае изменение наследственных свойств было связано с переносом ДНК.
Изучение химической структуры ДНК и генетических функций позволяет рассматривать гены как участки нуклеиновой кислоты, характеризующиеся определенной специфической последовательностью нуклеотидов. Расшифровка материальной сущности гена – одно из важных достижений современной биологической науки.
Линейное расположение генов в хромосомах. Сцепление генов. Кроссинговер
Основа принципа построения генетических карт Существование кроссинговера побудило Моргана разработать в 1911-1914 гг. принцип построения генетических карт хромосом.
В основу этого принципа положено представление о расположении генов по длине хромосомы в линейном порядке. За единицу расстояния между двумя генами условились принимать 1 % перекреста между ними.
Допустим, что к одной группе сцепления относятся гены А и В.
Между ними обнаружен перекрест в 10 %. Следовательно, гены А и В находятся на расстоянии 10 единиц. Допустим далее, что к этой же группе сцепления относится ген С.


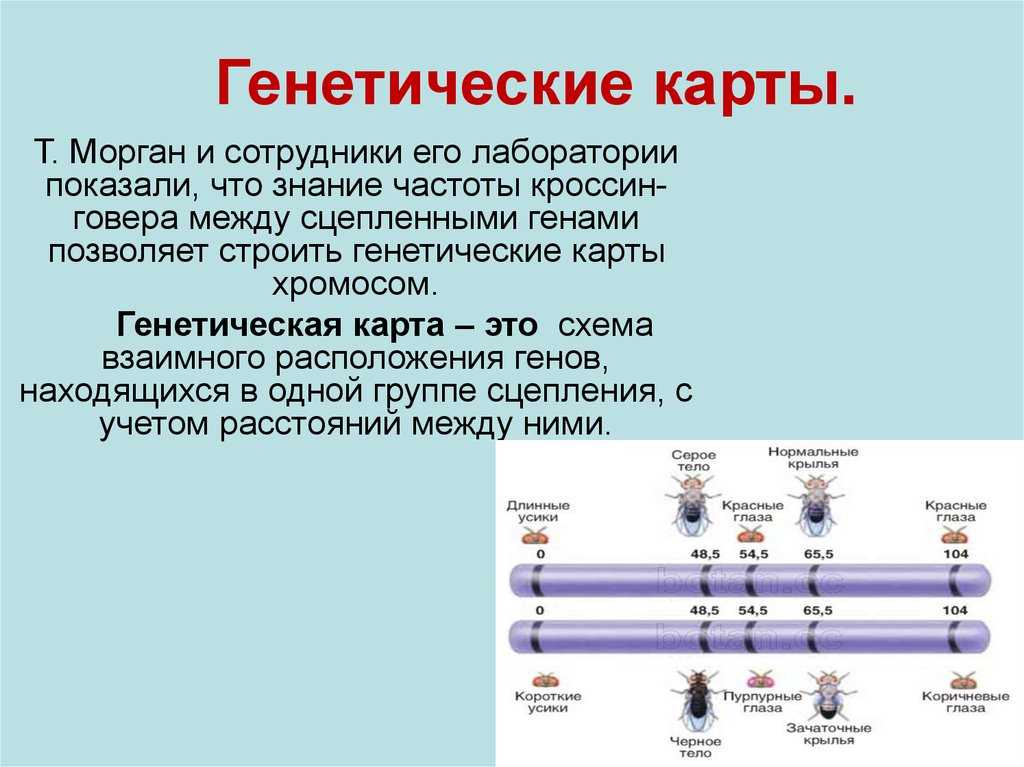

Чтобы узнать его место в хромосоме, необходимо выяснить, какой процент перекреста он дает с обоими из двух уже известных генов. Например, если с А он дает 3 % перекреста, то можно предположить, что ген С находится либо между А и В, либо с противоположной стороны, то есть А расположен между В и С.
Если между В и С окажется перекрест 7 %, то на хромосоме их следует расположить в таком порядке, как на верхней схеме. Если между В и С перекрест составит 13 %, то расположение генов будет как на нижней схеме.
Формула закономерности
В общей форме эту закономерность можно выразить следующей формулой: если гены А, В и С относятся к одной группе сцепления и расстояние между генами А и В равно нескольким единицам, а расстояние между В и С — одной единице, то расстояние между А и С может быть либо k +1, либо k-1.
СЦЕПЛЕНИЕ ГЕНОВ явление, в основе к-рого лежит локализация генов в одной хромосоме.Впервые обнаружено в 1906 У. Бэтсоном и Р. Пеннетом в опытах по скрещиванию душистого горошка.
Позднее С. г. было детально исследовано Т. Морганом с сотрудниками в экспериментах с дрозофилой. С. г. выражается в том, что аллели сцепленных генов, находящиеся в одной группе сцепления, имеют тенденцию наследоваться совместно. Это приводит к образованию у гибрида гамет преим. с «родительскими» сочетаниями аллелей.
Кроссинго́вер — процесс обмена участками гомологичных хромосом во времяконъюгации в профазе I мейоза. Помимо мейотического, описан также митотический кроссинговер. Хромосома разделяется на эти участки в определённых точках, одних и тех же для одного вида, что может быть определением вида на генетическом уровне, место расположение этих точек задаётся единственным геном.
Слайд 31При перицентрической инверсии, в случае перекреста между генами С и
Д, также получаются два продукта.Дупликация А и делеция F.Каждая из
полученных хромосом несет дупликацию одного неинвертированного района хромосом и делецию другого. В результате такие гаметы нежизнеспособны и кроссоверы не выявляются. Так же как и парацентрические, перицентрические инверсии «запирают» кроссинговер.
Поскольку кроссинговер в инвертированном участке хромосомы «заперт», в нем могут формироваться блоки мутаций, отличные от тех, которые локализованы в гомологичном фрагменте хромосомы, но не инвертированном. Это явление называют инверсионный полиморфизм популяций.

Определение пола[]
Основная статья: Определение пола
От чего же зависит рождение мужских и женских особей? Рассмотрим это на примере определения пола у дрозофилы. В ходе гаметогенеза у самок образуется один тип гамет, содержащий гаплоидный набор аутосом и одну X-хромосому. Самцы образуют два типа гамет, половина из которых содержит три аутосомы и одну X-хромосому (ЗА+Х), а половина — три аутосомы и одну У-хромосому (ЗА+У). При оплодотворении яйцеклеток (ЗА+Х) сперматозоидами с X-хромосомами будут формироваться самки (6А+ХХ), а от слияния яйцеклеток со сперматозоидами, несущими У-хромосому, — самцы (6A+XY). Поскольку число мужских гамет с X- и У-хромосомами одинаково, то и количество самцов и самок тоже одинаково. В данном случае пол организма определяется в момент оплодотворения и зависит от хромосомного набора зиготы.
Сходный способ определения пола (XY-тип) присущ всем млекопитающим, в том числе и человеку, клетки которого содержат 44 аутосомы и две X-хромосомы у женщин либо XY-хромосомы у мужчин.
Таким образом, XY-тип определения пола, или тип дрозофилы и человека, — самый распространенный способ определения пола, характерный для большинства позвоночных и некоторых беспозвоночных. Х0-тип встречается у большинства прямокрылых, клопов, жуков, пауков, у которых Y-хромосомы нет вовсе, так что самец имеет генотип Х0, а самка — XX.
У всех птиц, большинства бабочек и некоторых пресмыкающихся самцы являются гомогаметным полом, а самки —- гетерогаметным (типа XY или типа ХО). Половые хромосомы у этих видов обозначают буквами Z и W, чтобы выделить таким образом данный способ определения пола; при этом набор хромосом самцов обозначают символом ZZ, а самки — символом ZW или Z0.
Доказательства того, что половые хромосомы определяют пол организма, были получены при изучении нерасхождения половых хромосом у дрозофилы. Если в одну из гамет попадут обе половые хромосом, а в другую — ни одной, то при слиянии таких гамет с нормальными могут получиться особи с набором половых хромосом ХХХ, ХО, ХХУ и др. Выяснилось, что у дрозофилы особи с набором ХО — самцы, а с набором ХХУ — самки (у человека — наоборот). Особи с набором ХХХ имеют гипертрофированные признаки женского пола (сверхсамки). (Особи со всеми этими хромосомными аберрациями у дрозофилы стерильны). В дальнейшем было доказано, что у дрозофилы пол определяется соотношением (балансом) между числом X-хромосом и числом наборов аутосом.
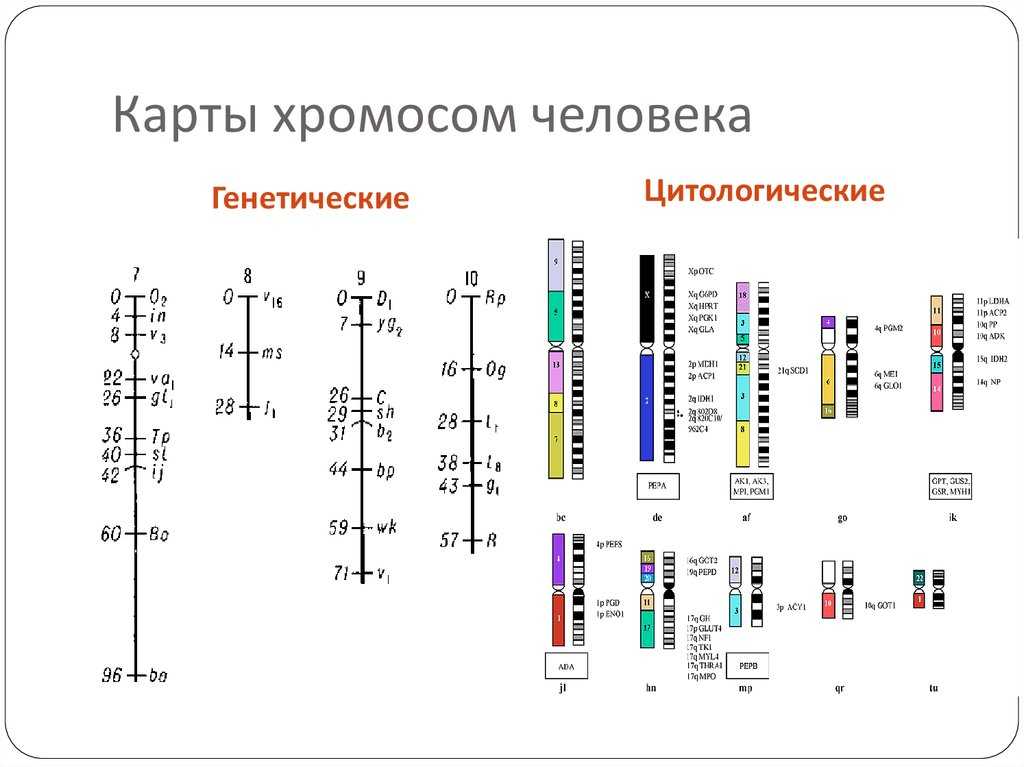
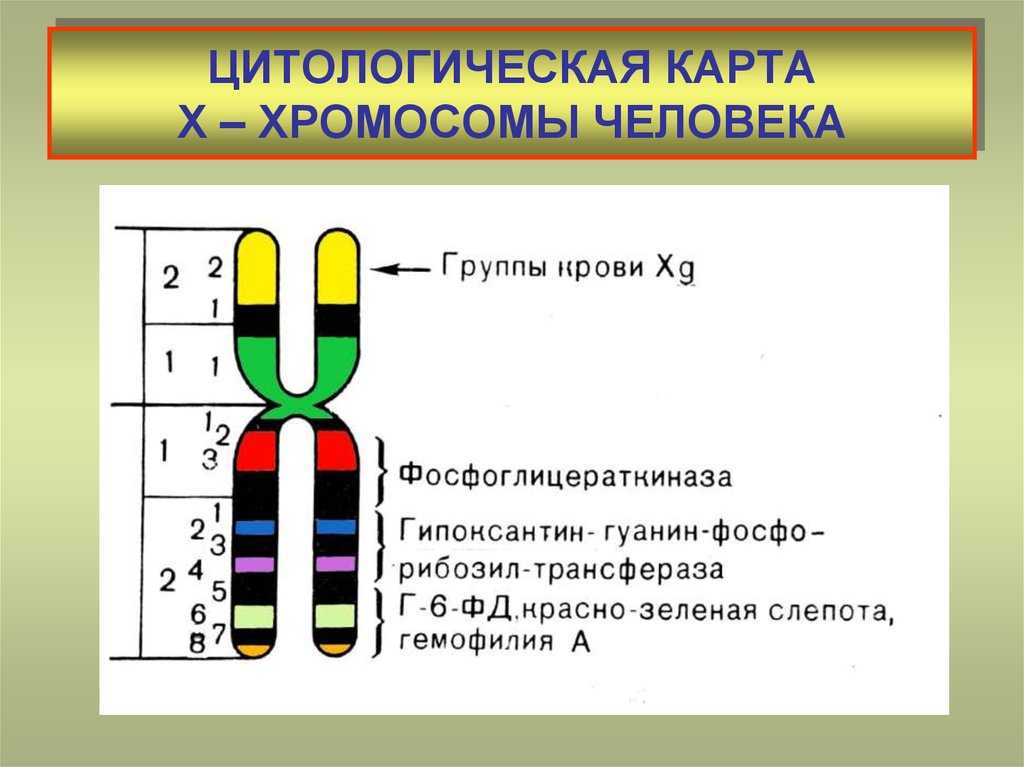
Что такое цитологические карты хромосом и как они составляются?
Если определить локализацию генов какого-либо организма непосредственно на хромосоме, то тем самым будет создана цитологическая карта данной хромосомы. Цитологическая карта хромосомы, в которой можно наблюдать истинное расположение генов, впервые создана для дрозофилы и кукурузы. Метод создания цитологических карт заключается в следующем: с помощью мутагенов индуцируют различные хромосомные перестройки — делеции, транслокации или инверсии, которые приводят к изменению силы сцепления генов, что выявляется при генетическом анализе. Некоторые из этих перестроек можно определить с помощью микроскопа и установить корреляцию между генетическими и цитологическими данными.
Так, если определенное количество генов, принадлежащих к одной группе сцепления, в результате транслокации окажется в другой, то можно измерить величину транслоцированного фрагмента в световом микроскопе и составить его цитологическую карту. В мейозе при конъюгации с хромосомой, несущей делецию, нормальная хромосома образует петлю напротив того места, где в гомологичной хромосоме произошла делеция, что дает возможность определить локализацию выпавшего фрагмента геном В. Фенотипически такое изменение может проявиться в «ложном доминировании» A CD/abcd признака, обусловленного рецессивным геном b. После сопоставления цитологических и генетических данных можно сделать вывод, что ген В локализован в выпавшем фрагменте, положение которого в хромосоме установлено.
При составлении цитологических карт учитываются также некоторые особенности слюнных желез личинок насекомых. Слюнные железы насекомых достигают больших размеров без какого бы то ни было деления клеток или ядер. При этом хромосомы клеток слюнных желез раскручиваются и становятся длиннее хромосомы обычных клеток. Затем хромосомы многократно удваиваются, но дочерние хромосомы не расходятся. Такое явление называется эндомитозом. В результате образуются длинные лентовидные хромосомы с хорошо заметными поперечными полосами по всей длине, т. е. политенные хромосомы (рис. 15).
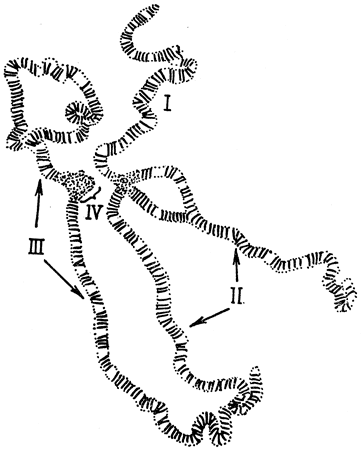 Рис. 15. Политенные хромосомы дрозофилы. Цифрами указаны группы сцепления
Рис. 15. Политенные хромосомы дрозофилы. Цифрами указаны группы сцепления
Полосы или диски хорошо окрашиваются. Поперечная исчерченность для каждой хромосомы имеет характерный вид. Число и положение дисков специфично для каждой хромосомы. Диски служат маркерами. С их помощью можно установить характер и размеры хромосомных перестроек, а также положение генов, существование которых установлено по данным рекомбинационного анализа. Например, делецию, величина которой найдена при помощи генетического анализа, можно связать с исчезновением определенной полосы или группы полос на такой гигантской хромосоме. Когда цитологическую карту сравнивают с соответствующей генетической картой, линейный порядок расположения генов полностью совпадает, однако могут иметь место расхождения в относительном положении (расстоянии) генов. Так, например, вблизи центромеры гены, которые на генетической карте кажутся тесно сцепленными, на самом деле расположены значительно дальше друг от друга. Это несовпадение генетических и цитологических карт объясняется неравномерной частотой перекреста по длине хромосомы при кроссинговере.
Сопоставление генетических и цитологических карт дало возможность подтвердить следующие положения хромосомной теории наследственности:
1) хромосомы по своей длине наследственно дискретны;
2) каждый ген имеет определенное место (локус) в хромосоме;
3) гены расположены в хромосоме в определенной линейной последовательности;
4) частота кроссинговера между генами зависит от расстояния между ними.
Сцепленное наследование[]
Независимое комбинирование признаков (третий закон Менделя) осуществляется при условии, что гены, определяющие эти признаки, находятся в разных парах гомологичных хромосом. Следовательно, у каждого организма число генов, способных независимо комбинироваться в мейозе, ограничено числом хромосом. Однако в организме число генов значительно превышает количество хромосом. Например, у кукурузы до эры молекулярной биологии было изучено более 500 генов, у мухи дрозофилы — более 1 тыс., а у человека — около 2 тыс. генов, тогда как хромосом у них 10, 4 и 23 пары соответственно. То, что число генов у высших организмов составляет несколько тысяч, было ясно уже У. Сэттону в начале XX века. Это дало основание предположить, что в каждой хромосоме локализовано множество генов. Гены, локализованные в одной хромосоме, образуют группу сцепления и наследуются вместе.
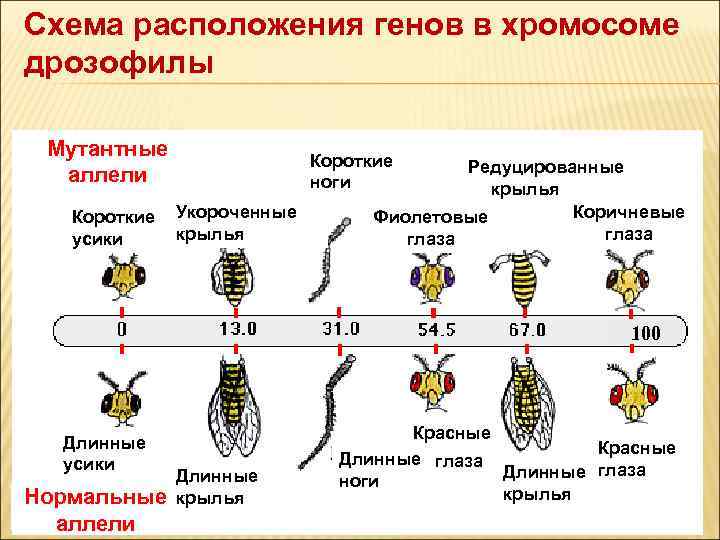
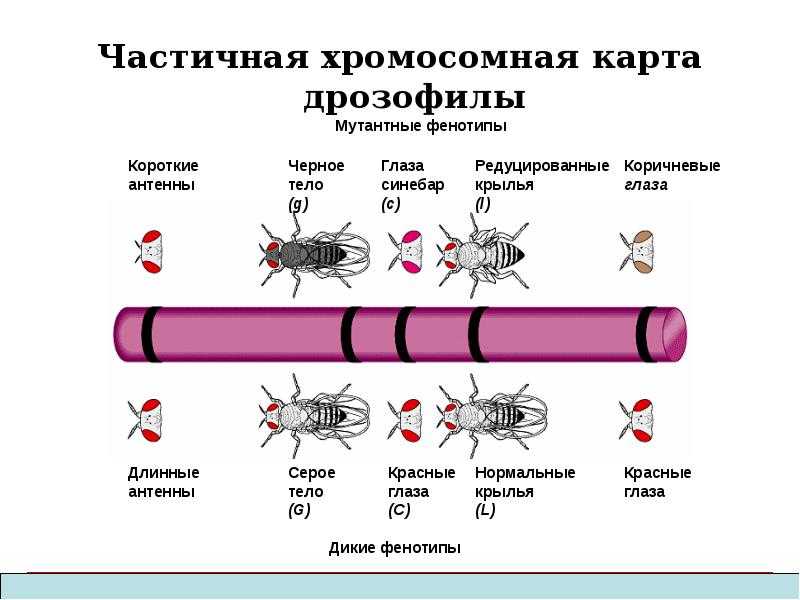


Совместное наследование генов Т. Морган предложил назвать сцепленным наследованием. Число групп сцепления соответствует гаплоидному числу хромосом, поскольку группу сцепления составляют две гомологичные хромосомы, в которых локализованы одинаковые гены. (У особей гетерогаметного пола, например у самцов млекопитающих, групп сцепления на самом деле на одну больше, так как X- и У-хромосомы содержат разные гены и представляют собой две разные группы сцепления. Таким образом, у женщин 23 группы сцепления, а у мужчин — 24).
Способ наследования сцепленных генов отличается от наследования генов, локализованных в разных парах гомологичных хромосом. Так, если при независимом комбинировании дигетерозиготная особь образует четыре типа гамет (АВ, Ab, аВ и ab) в равных количествах, то при сцепленном наследовании (в отсутствие кроссинговера) такая же дигетерозигота образует только два типа гамет: (АВ и ab) тоже в равных количествах. Последние повторяют комбинацию генов в хромосоме родителя.
Было установлено, однако, что кроме обычных (некроссоверных) гамет возникают и другие (кроссоверные) гаметы с новыми комбинациями генов — Ab и аВ, отличающимися от комбинаций генов в хромосомах родителя. Причиной возникновения таких гамет является обмен участками гомологичных хромосом, или кроссинговер.
Кроссинговер происходит в профазе I мейоза во время конъюгации гомологичных хромосом. В это время части двух хромосом могут перекрещиваться и обмениваться своими участками. В результате возникают качественно новые хромосомы, содержащие участки (гены) как материнских, так и отцовских хромосом. Особи, которые получаются из таких гамет с новым сочетанием аллелей, получили название кроссинговерных или рекомбинантных.
Частота (процент) перекреста между двумя генами, расположенными в одной хромосоме, пропорциональна расстоянию между ними. Кроссинговер между двумя генами происходит тем реже, чем ближе друг к другу они расположены. По мере увеличения расстояния между генами все более возрастает вероятность того, что кроссинговер разведет их по двум разным гомологичным хромосомам.
Расстояние между генами характеризует силу их сцепления. Имеются гены с высоким процентом сцепления и такие, где сцепление почти не обнаруживается. Однако при сцепленном наследовании максимальная частота кроссинговера не превышает 50 %. Если же она выше, то наблюдается свободное комбинирование между парами аллелей, не отличимое от независимого наследования.
Биологическое значение кроссинговера чрезвычайно велико, поскольку генетическая рекомбинация позволяет создавать новые, ранее не существовавшие комбинации генов и тем самым повышать наследственную изменчивость, которая дает широкие возможности адаптации организма в различных условиях среды. Человек специально проводит гибридизацию с целью получения необходимых вариантов комбинаций для использования в селекционной работе.
Слайд 39При дупликации, наоборот, можно ожидать увеличение на 50 % активности
ферментов, гены которых вовлечены в дупликацию. Самым известным примером картирования
гена с помощью дупликации является супероксиддисмутаза, которая была картирована на хромосоме 21, так как ее уровень был постоянно повышен у больных с болезнью Дауна.
Исследованы геныAPP: предшественник протеина бета-амилоида (A4) (пептидаза нексин-II, болезнь Альцгеймера) CBS: цистатионин бета-синтаза CLDN14: Клаудина 14 HLCS: голокарбоксилаза синтетаза (биотин- (проприонил-коекзим A-карбоксилаза (ATP-гидролитических) лигаза) KCNE1: потенциал управляемый калиевый канал, член 1, семья Isk-связанных (англ. Potassium voltage-gated channel, Isk-related family, member 1) KCNE2: потенциал управляемый Калыев канал, член 2, семья Isk-связанных (англ. Potassium voltage-gated channel, Isk-related family, member 2) LAD: дефицит адгезии лейкоцитов (возможно также обозначения ITGB2, CD18, LCAMB) SOD1: супероксиддисмутаза 1, растворимый (амиотрофический боковой склероз 1 (у взрослых)) TMPRSS3: трансмембранным протеазы, серин 3 PCNT: центросомний перицентрин DSCR1: критическая участок 1 синдромом Дауна DYRK1A: киназа 1A, регулирующего тирозин- (Y) -фосфориляцию с двойной специфичностью RRP1B: Рибосомальные процессинг РНК 1 гомолог B S100B: кальций-связывающий белок из семейства S100
