

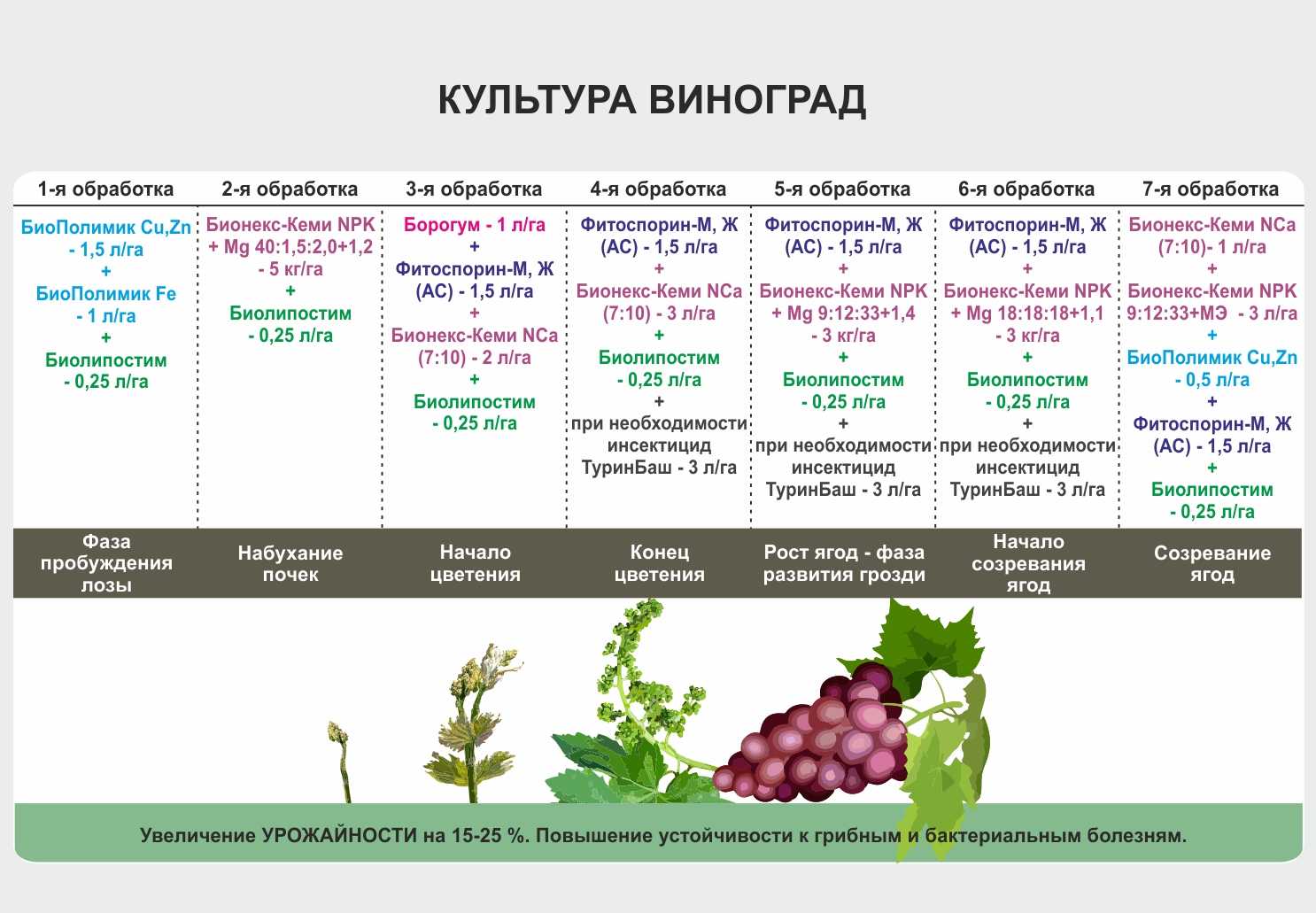
Особенности селекции животных
Селекция животных — это область науки, изучающая наиболее оптимальные способы выведения пород домашних животных и улучшения существующих пород.
В селекции животных используют все методы селекции, но эти методы имеют свою специфику, связанную с отсутствием у домашних животных способности к бесполому и вегетативному размножению, а также с особенностями получения потомства — у домашних животных достаточно поздно наступает период половой зрелости и потомство относительно немногочисленно.
При выведении новых пород животных большее применение имеет индивидуальный отбор, так как за животными осуществляется более индивидуальный уход, чем за растениями (например, за стадом в 100 голов ухаживает до 10 человек, в то время как поле, на котором произрастают сотни тысяч растений, обслуживает бригада в 5-8 человек).
Важное значение в селекции животных имеет применение гибридизации, при этом используют инбридинг, неродственное скрещивание и отдаленную гибридизацию. Неродственное скрещивание представляет собой гибридизацию животных, принадлежащих к разным породам одного вида
Неродственное скрещивание представляет собой гибридизацию животных, принадлежащих к разным породам одного вида.
Такое скрещивание приводит к «расшатыванию» наследственности и получению организмов с новыми признаками, которые можно в дальнейшем использовать для выведения новой породы или улучшения старой.
Инбридинг у животных применяется в целях, аналогичных инбридингу у растений, только в отличие от самоопыления растений здесь используют осеменение близкородственных организмов (матери, сестры, дочери и т. д.).
Отдаленная гибридизация проводится для получения животных с ценными эксплуатационными свойствами (так скрещивают осла с лошадью, яка с туром). Полученные гибриды, как правило, потомства не дают.
Важную роль играет направленное воспитание, позволяющее развить и закрепить полезное для хозяйственной деятельности свойство животного.
Классическим примером селекции животных является выведение М. Ф. Ивановым белой украинской породы свиней. Эта порода характеризуется высокой продуктивностью и хорошей приспособленностью к местным условиям. Для выведения этой породы использовали местную украинскую породу, хорошо приспособленную к условиям существования в степях, но обладавшую малой продуктивностью и невысоким качеством мяса. Другой породой была английская белая порода, характеризующаяся высокой продуктивностью, но отсутствием приспособленности к существованию в условиях Украины. Были использованы инбридинг, неродственное скрещивание, индивидуально-массовый отбор, воспитание условиями содержания. Длительная и кропотливая работа дала положительный результат — была выведена новая порода свиней — украинская белая свинья, сочетающая в себе качества обоих родительских пород в положительном для хозяйственной деятельности контексте — высокая продуктивность и хорошая приспособленность к местным условиям.
М.Ф.Ивановым с коллегами была проведена большая работа по отдаленной гибридизации, в результате которой выведены архаромериносы (гибрид горного барана — архара и овцы-мериноса), зубробизоны и т. д.
Ограниченная передача генов пластид между привоем и подвоем
В 2009 году в журнале Science была опубликована статья Ральфа Бока (англ. Ralph Bock) и Сандры Стегеманн (англ. Sandra Stegemann), в которой доказана возможность обмена генами между привоем и подвоем двух трансгенных линий табака, несущих хлоропластные и ядерные маркеры. Верхушку растения одной из линий использовали в качестве привоя. После срастания привоя и подвоя участок растения, прилежащий к зоне срастания, вырезали и получали из него культуру клеток. В 45 случаях из 74 из вырезанного участка ткани были получены клеточные линии, содержащие оба маркера (ядерный маркер одного из сортов и хлоропластный маркер другого сорта). Авторы доказали, что происходила передача генов хлоропластов (либо больших участков хлоропластного генома, либо целых пластид) в обоих направлениях (от привоя к подвою и наоборот). Передачи ядерных генов не наблюдали ни в одном из случаев. Показано также, что это явление не связано со слиянием клеток привоя и подвоя и с разрезанием тканей растений. Из клеточных линий, содержащих оба маркера, были выращены взрослые растения. Из их семян выросли растения с тем же признаком (двойной устойчивостью к антибиотикам), что и у «гибридных» клеток, образовавшихся на границе привоя и подвоя. Таким образом, новая комбинация признаков, полученная в результате «вегетативной гибридизации», действительно может стать наследуемой, по крайней мере в лабораторных условиях.
Однако при этом авторы статьи отмечают, что хотя эти «данные демонстрируют обмен генетическим материалом между прививаемыми растениями, они не поддерживают догмат лысенкоизма о том, что „прививочная“ гибридизация была бы аналогичной гибридизации путём полового процесса». Вместо этого «открытие того, что перенос генов ограничен зоной контакта между привоем и подвоем, указывает на то, что данные изменения могут наследоваться только путём формирования бокового побега из зоны прививки». Все другие побеги будут содержать только гены одного из исходных растений. Более того, неизвестно, могут ли подобные процессы происходить в природе. «Ясно лишь, что это должно происходить гораздо реже, чем предполагали сторонники лысенкоизма». Кроме того, «вопреки взглядам лысенкоистов, „вегетативная гибридизация“ влияет только на крайне ограниченное число признаков, определяющихся геномами органелл — пластид и митохондрий», но не ядерными генами.
Молекулярный биолог Николь Раск (англ. Nicole Rusk) так пишет о работе Ральфа Бока и Сандры Стегеманн: «Демонстрируя данные по переносу генов при прививках лишь на ограниченное расстояние, Бок представляет также молекулярные данные о том, что концепция вегетативной гибридизации является нежизнеспособной… Сегодня догмат Лысенко в значительной степени дискредитирован, хотя существует несколько опубликованных сообщений, заявляющих о наблюдениях наследуемых изменений в растении после процедуры прививки. Согласно Боку, эти сообщения поступают, в основном, от растениеводов, но молекулярные данные в пользу вегетативной гибридизации в настоящее время отсутствуют».
Биология — Вегетативная гибридизация
С древнейших времён до XIX века Прививочные гибриды в работах Дарвина Работы Винклера и Баура Прививочные гибриды в работе селекционеров-практиков Дискуссия о вегетативной гибридизации в СССР Взгляды современных последователей Лысенко и попытки реабилитации «вегетативной гибридизации» Химеры растений и прививочные гибриды Ограниченная передача генов пластид между привоем и подвоем
Схема прививки почки: 1 почка привоя удаляется вместе с подлежащими тканями; 2-4 почка вставляется в Т-образный разрез на стебле подвоя и там закрепляется, 5 почка образует побег
Вегетативная гибридизация — постулируемое в рамках «мичуринской агробиологии» Т. Д. Лысенко и его последователей явление образования при прививках растений «вегетативных гибридов», сохраняющих при половом размножении модификации, приобретенные в результате прививок, и не отличающихся от истинных гибридов, получаемых скрещиванием. «Вегетативная гибридизация» служила для последователей Лысенко одним из центральных «доказательств» ложности классической генетики и хромосомной теории наследственности.








Понятие о вегетативных гибридах было введено И. В. Мичуриным, который использовал его вместо принятого к тому времени термина «прививочный гибрид». Первоначально вегетативной гибридизацией называли изменения привоя под влиянием подвоя при прививках растений. Позже Лысенко и его последователи объединили под именем вегетативной гибридизации довольно разнородные явления, связанные, по их представлениям, с «расшатыванием» наследственности; они не только рекомендовали вегетативную гибридизацию как важный прием практической селекции, но и использовали данные по вегетативной гибридизации как аргумент, призванный опровергнуть хромосомную теорию наследственности и подтвердить их ламаркистские воззрения.
В результате серии экспериментальных исследований 1930—1950-х годов наличие вегетативной гибридизации подтвердить не удалось, а развитие генетики во второй половине XX века показало ложность теоретических воззрений школы Лысенко. Поскольку образующийся в результате прививки или трансплантации организм относится не к истинным гибридам, а к химерам, в современной научной литературе термины «вегетативный гибрид» и «вегетативная гибридизация» практически не используются.
В учебнике Лобашева термин «вегетативная гибридизация» рассматривается как эквивалент термина «прививочная гибридизация», а сам вопрос о влиянии подвоя на привой рассматривается в духе классической генетики. Более современные университетские учебники по генетике либо содержат жесткую критику теории вегетативной гибридизации, причем само название признается неудачной заменой более раннего термина «прививочный гибрид», которое тоже рекомендовано использовать лишь как иносказание, либо вообще не упоминают ни о «прививочных», ни о «вегетативных» гибридах. В монографии А. Е. Гайсиновича по истории генетики понятие «вегетативной гибридизации» используется только при описании работ Лысенко и его сторонников.
|
Кругосветное путешествие Чарльза Дарвина |
Вейсманизм-морганизм >>> |
Вегетативная гибридизация
Учение о вегетативной гибридизации как методе практической селекции создал И.В. Мичурин Им был разработан метод ментора (воспитателя), который основан на взаимном влиянии подвоя и привоя.
И.В. Мичурин доказал, что сорта плодовых культур, представляющие собой уже сформировавшиеся организмы, не могут изменяться под влиянием прививки. Поэтому, например, при прививке старых сортов
винограда к старым сортам филлоксероустойчивых подвоев или к морозоустойчивому амурскому винограду не наблюдается изменения наследственных свойств ни привоя, ни подвоя.
Изменение наследственных свойств при прививке происходит только у молодых гибридных организмов с расшатанной наследственностью. Путем прививки черенков гибридных сеянцев к старым со сложившейся наследственностью растениям можно направленно изменять свойства сеянцев.
Старые, сформировавшиеся растения (ментор) могут быть взяты и в качестве привоя. В этом случае, прививая к сеянцу в разные периоды его развития черенок определенного сорта или вида и выдерживая определенную продолжительность действия ментора, селекционер воспитывает сеянец в нужном направлении; он усиливает и развивает в нем хорошие качества и задерживает, а иногда и совершенно уничтожает склонность к проявлению нежелательных свойств.
Происходящие в этом случае изменения наследственных свойств обусловлены обменом веществ, происходящим между подвоем и привоем, взаимной ассимиляцией веществ, вырабатываемых каждым из компонентов.
Применяя метод ментора, И.В. Мичурин получил ряд сортов плодовых культур.
Метод ментора у винограда, так же как и у плодовых культур, следует применять путем:
1) прививки гибридного сеянца на старый подвой (с удалением надземной его части) для усиления роста слабо растущего сеянца и придания ему нужных свойств – укороченного периода вегетации и др.;
2) прививки сеянца в крону старого сорта для питания сеянца за счет подвоя; в этом случае часть листьев на побеге сеянца удаляют; этим способом можно значительно изменить свойства сеянца в отношении его морозоустойчивости, устойчивости против болезней, качества плодов и т.д.;.
3) прививки черенка старого сорта к сеянцу; этот способ применяется на более поздних стадиях развития сеянца для улучшения качества плодов и других целей.
В виноградарстве широко применяются способы прививки глазком, черенком в период покоя и в зеленом состоянии. Для вегетативной гибридизации винограда наиболее интересен способ прививки проростком сеянца за кору побега или способ зеленой прививки в расщеп сеянцем в семядольном состоянии (рис. 187).
Применяя вегетативную гибридизацию, виноградари должны постоянно учитывать опыт И.В. Мичурина и помнить его предупреждение, что неправильный выбор «воспитателя» может привести к неудаче.
При выращивании и воспитании сеянцев винограда вегетативная гибридизация применяется и для того, чтобы ускорить вступление их в пору плодоношения и получить такие формы, которые быстро и обильно плодоносят.
Сеянцы, в зависимости от наследственных свойств и условий их выращивания, начинают закладывать плодовые почки иногда в первый год, а иногда это происходит через 20 лет. Прививкой на стадийно старые, быстро вступающие в плодоношение сорта, а также различными хирургическими приемами (формировка, обрезка, прищипка и т.д.) можно значительно ускорить время плодоношения сеянца.
Задача селекции
Селекция винограда позволяет совершенствовать базовые сорта, а также, скрещивая их, создавать новые виноградные виды. Благодаря тому, что отбор проводится искусственно (человеком), можно выводить сорта, которые соответствуют местам их будущего выращивания. В процессе селективных работ из растения исключаются слабые характеристики.










Из-за этого виноградные кусты становятся более устойчивыми к внешним негативным факторам, заболеваниям, а также повышается качество урожая. Селекция позволяет создавать уникальные вкусы ягод, путем скрещивания между собой необычных виноградных сортов. Эта задача особенно важна в виноделии, так как такой виноград позволяет производить неповторимые напитки.
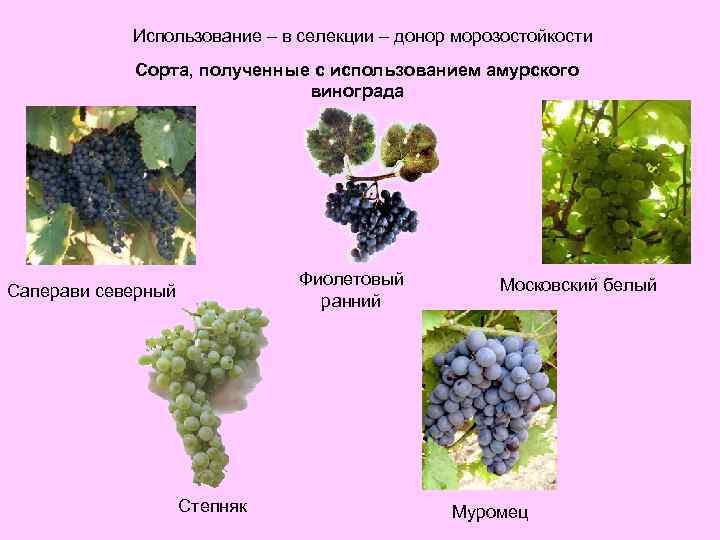
Благодаря селекционным работам появились морозоустойчивые сорта винограда.
Популярные представители
Среди сортов винограда, выведенных благодаря селекционном работам, можно выделить определённых представителей. По их характерным чертам возможно судить о возможностях селекции.
Морозостойкие
Одним из главных направлений в селекционных работах с виноградом является попытка увеличить его морозоустойчивость. Из-за того, что по своей природе виноград весьма теплолюбивое растение, его крайне сложно выращивать в северных регионах. Однако благодаря селекции были выведены сорта, способные противостоять пониженным температурам.
Сорт винограда Амурский прорыв – морозостойкий гибрид, выдерживает температуру до –40 °С.
Мариновский
Виноград среднего срока созревания, способен переносить понижение температуры до -30°C. При правильной агротехнике грозди могут достигать веса до 1 кг. В основном сорт используется для свежего потребления и изготовления сладких соков. Также из данного Мариновского сорта можно готовить домашние розовые и белые сухие вина.
Альфа
Технический сорт Альфа винограда северо-американского происхождения. Идеальный вариант для средней полосы России. Способен переносить морозы до -35°C над землей, а корневая система выдерживает до -12°C. Ягоды по вкусу напоминают сорт Изабелла.
Сорт винограда Alfa (Альфа) – технический неукрывной сорт черного винограда, выдерживает морозы до –40 °С.
Элегант
Виноград раннего созревания с высокими вкусовыми качествами. Является столовым сортом, поэтому подходит для изготовления соков, джема, варенья, а также употребления в свежем виде. Элегант хорошо переносит жару и засуху, а зимой способен выстоять при температуре -25°C.
Болезнеустойчивые
Не менее важная проблема при выращивании винограда – устойчивость к заболеваниям. Из-за того, что большая часть России не является изначально подходящей территорией для выращивания этой культуры, кусты могут быть более восприимчивы к болезням. Благодаря селекции эта проблема постепенно решается.
Тимур
Столовый сорт винограда с ранним сроком созревания. Бывает двух видов – белый и розовый. Оба вида сорта Тимур устойчивы к милдью и оидиуму, а также спокойно переносят снижение температуры до -24°C.
Сорт винограда Тимур разделяется на два вида, кроме белого есть ещё и розовый Тимур.
Венус
Бессемянный сорт раннего созревания. Виноград отличается высокой урожайностью и отсутствием семян. Устойчив к оидиуму и милдью , но при влажной погоде зрелые грозди могут поражаться серой гнилью. Морозостойкий – выдерживает до -26°С. Используется при приготовлении вина, соков, а также употребляется в свежем виде.
Гарольд
Белый виноград сверхраннего созревания. Гарольд может давать двойной урожай за счет плодоношения побегов и пасынков. Высокая устойчивость к серой гнили, милдью и оидиуму. Зимой способен выдерживать понижение температуры до -25°С.
Химеры растений и прививочные гибриды
Согласно современным представлениям, взаимное влияние привоя и подвоя существует, однако является следствием объединения в физиологически единый организм генетически разнородных частей, то есть образование химеры различных типов. Химеры подробно обсуждаются в монографии Н. П. Кренке (1947). Современная трактовка содержится в более поздних публикациях.
При этом на формирование признаков компонентов химеры оказывает влияние транспорт метаболитов, выделяемых фрагментами (в том числе и фитогормонов). Однако поскольку генеративные органы химеры обычно формируются только подвоем, а в половом процессе участвуют гаметы, образующиеся только из одной из клеточных линий химеры, обмен генетическим материалом не происходит и при размножении семенами признаки гибрида не наследуются. Типичным примером является ракитник Адама, семена которого дают исходный желтоцветный ракитник.
В случае пестролистных химер также может проявляться и пластидное наследование, описанное Бауром и Корренсом в начале XX века: так, в случае пестролистной ночной красавицы Mirabilis jalapa пластиды, синтезирующие хлорофилл, передаются с цитоплазмой яйцеклетки: так, при опылении цветков на белых (бесхлорофильных) побегах пыльцой цветков любых форм (белые, зелёные, пёстрые) из гибридных семян всегда вырастают белые, лишённые хлорофилла и вследствие этого нежизнеспособные растения; при опылении цветков на зелёных побегах картина противоположна (только зелёные); при опылении цветков на пестрых побегах гибридные семена дают все три формы.
Это явление служит подтверждением образования гамет из одной клеточной линии химеры и отсутствия внегенетической передачи признаков.
В современной литературе для химер, полученных прививкой, используется термин «прививочные гибриды». Взаимное влияние привоя и подвоя используется для усиления фенотипического проявления желаемых признаков, то есть модификаций, сохраняющихся при вегетативном размножении.










2020-10-01 06:47:41:Вегетативная гибридизация
Вегетативная гибридизация
Идея вегетативной гибридизации была дискредитирована попытками Лысенко с её помощью отрицать законы генетики и хромосомную теорию, хотя многочисленные попытки не давали результата. Современные исследования показывают не только возможность переноса целых геномов, но и их полное соответствие генетическим законам.
Проблема межвидового скрещивания заключается в невозможности схождения гомологичных хромосом из-за разницы кариотипов. Однако растения проще переносят полиплоидизацию, что позволяет создавать новые гибридные виды растений с помощью горизонтального переноса ядерных геномов. Аллополиплоидизация вероятно лежит в основе формирования многих современных диких и культурных растений. Но данный процесс весьма затруднён, так как требует предварительного межвидового скрещивания, т.е. слияния двух половых клеток с образованием аллодиплоидной зиготы. А это имеет ряд трудностей, таких как межвидовой барьер и стерильность образовавшихся гибридов. В естественных условиях гибриды могут сформироваться в полноценный новый вид путём вегетативного размножения до тех пор, пока не произойдёт случайная полногеномная дупликация. В это случае решается проблема формирования гомологичных пар хромосом, а значит аллотетраплоидный гибрид восстанавливает способность к половому размножению и может считаться полноценным новым видом, генетически изолированным от родительских.
Другим путём видообразования можно представить, объединение ядерных геномов диплоидных вегетативных клеток, что сразу бы давало аллотетраплоидные гибриды способные к половому размножению. Но данный процесс в течение многих лет не находил экспериментального подтверждения, и многие специалисты склонялись к мысли о его невозможности. В 2009 году были опубликованы работы, показывающие обмен пластидными геномами меду клетками привоя и подвоя, что показало техническую возможность вегетативной гибридизации. Спустя пять лет Штегеманном и Боком с коллегами был доказан горизонтальный перенос и пластидных и ядерных геномов между клетками сросшихся растений. Далее были проведены работы по получению тетраплоидных гибридов обыкновенного табака и табачного дерева. При обычном скрещивании этих видов образуются бесплодные гибриды, а полученные в ходе эксперимента аллотетраплоиды оказались фертильны и показали более высокую скорость роста в сравнении с обоими родительскими видами.
Полученные результаты показывают не только не только перспективный метод получения новых культурных растений, но и раскрывают сущность одного из возможных естественных механизмов видообразования. Вегетативная гибридизация изначально служащая для опровержения хромосомной теории наследственности и законов генетики в итоге их полностью подтверждает.
Прививочные гибриды в работах Дарвина
В известном труде «Изменение животных и растений в домашнем состоянии» (англ. Charles Darwin «The Variation of Animals and Plants Under Domestication», 1868 год) Дарвин отвёл целый раздел этой проблеме, назвав его «Гибриды, происходящие вследствие прививки» (англ. Graft-hybrids). Дарвин описал известные ему по данным литературы случаи образования помесей между отдельными видами или разновидностями без участия полового размножения. В существовании таких гибридов он был «убеждён» и считал такие факты «чрезвычайно важными» и способными «рано или поздно изменить взгляды физиологов на половое воспроизведение». (англ. „most important fact, which will sooner or later change the views held by physiologists with respect to sexual reproduction“).
В большинстве рассматриваемых примеров Дарвин отметил неоднородность признаков тканей и органов прививочных гибридов с достаточно чёткими пространственными границами, склонность к возвращению частей со смешанными признаками к исходным родительским формам (реверсия). Однако, он считал, что «разделение подобного вида происходит отнюдь не редко … и у семенных гибридов» (англ. „But segregation of this kind occurs by no means rarely … in seminal hybrids“).
В качестве примеров прививочных гибридов Дарвин наряду с ракитником Адама рассматривал и Bizzarria (прививочный гибрид померанца с лимоном, известный с XVII века, для которого было характерно присутствие на одном растении как побегов и плодов исходных форм, так и «гибридных» плодов разнообразных форм: и со смешением признаков померанца и лимона, и состоявших из чётко различимых фрагментов, образованных тканями лимона и померанца), отметив, что Bizzarria размножается только черенками.
Дарвин рассмотрел различные гипотезы, объяснявшие возникновение и возможные причины вариабельности прививочных гибридов в рамках представлений о сохранении видовой (или сортовой) индивидуальности привоя и подвоя: изменение потока питательных веществ при прививке на подвой другого вида, возможное взаимное инфицирование привоя и подвоя, спонтанная изменчивости почек, ошибочно рассматриваемая как образование прививочного гибрида.
Подводя итог, Дарвин отметил, что во многих случаях — например, при прививках плодовых деревьев — наблюдаемое смешение признаков нельзя рассматривать как гибридизацию, но иногда — в частности, в опытах Магнуса с картофелем, случаи получения промежуточных форм при прививках «кажется, доказывают, что при некоторых неизвестных условиях гибридизация путём прививки возможна» (англ. Nevertheless, the cases above given seem to me to prove that under certain unknown conditions graft-hybridisation can be effected) и высказал предположение, что некие «элементы, которые идут на образование нового существа» могут содержаться не только в половых органах, но и в соматической ткани.
Явления изменчивости при прививках Дарвин рассматривал лишь как один из штрихов общей картины разнообразия форм наследования и изменчивости одомашниваемых видов и проблемы соотношения передачи признаков при половом и вегетативном размножении. Результатом стало сделанное Дарвином в главе XXVII «Временная гипотеза пангенезиса» (англ. Provisional hypothesis of pangenesis) предположение, что во всех тканях организмов присутствуют субмикроскопические гранулы — геммулы, несущие наследственные признаки. Перенос геммул в репродуктивные органы, согласно этой гипотезе, — универсальный механизм, объясняющий весь комплекс наследственности и изменчивости. Дарвин предупреждал, что пангенез — не более, чем временная умозрительная гипотеза; вскоре после опубликования дарвиновской работы — в 1871 году в серии экспериментов по проверке существования геммул, поставленной Фрэнсисом Гальтоном (двоюродный брат Ч. Дарвина), гипотеза не подтвердилась. В этих опытах Ф. Гальтон переливал кровь от темноокрашенных кроликов светлоокрашенным, и никакого влияния на окраску шерсти у потомства не обнаружил.