

Определение бактериальной трансдукции – что такое трансдукция?
- Трансдукция относится к переносу части ДНК между двумя бактериями с помощью бактериофага.
- Трансдукция осуществляется вирусами.
- При репликации вируса в клетке небольшой фрагмент бактериальной ДНК встраивается в бактериофаг. Затем эта ДНК передается бактериям-реципиентам во время заражения.
- Благодаря известному процессу лизогенной конверсии ДНК фага реципиента бактериальная клетка встраивается в клеточную ДНК.
- Лизогенная трансформация придает новое свойство бактериальная клетка. Например, непатогенные бактерии могут стать патогенными в результате лизогенной конверсии.
- Бактериофаги могут кодировать дифтерийный, ботулинический, холерный и эритрогенный токсины одной бактерии посредством трансдукции.
- Бактерии, заражающие вирусы, известны под аббревиатурой бактериофаги (или фаги).
- Вирулентные бактериофаги размножаются внутри своего бактериального хозяина, как только они проникают внутрь. После того, как частицы дочернего фага достигают определенного количества, они вызывают лизис клеток-хозяев. Это позволяет им высвобождаться и заражать другие клетки-хозяева. Этот процесс называется «литическим циклом».
- Однако умеренные бактериофаги не сразу убивают своего хозяина. Вместо этого хозяин устанавливает с фагом отношения, называемые лизогенией. Лизогенизированные бактерии также известны как лизогены.
- Многие виды фагов умеренных широт устанавливают лизогению путем встраивания своего генома в бактериальную клетку. хромосома. Профаги — это вирусный геном, который встраивается в бактерию-хозяина. Это не вредно для бактерии-хозяина, и ген фага копируется вместе с геномом клетки-хозяина.
- Умеренные фаги могут оставаться неактивными в своих хозяевах в течение нескольких поколений. При определенных условиях, таких как УФ-излучение, их можно заставить переключиться на литический цикл. Как только это происходит, профаг из генома бактерий удаляется, и литические циклы продолжаются.
Неспецифическая, или общая, трансдукция
Для неспецифической Трансдукции не требуется состояния лизогении с внедрением профага вблизи тех бактериальных генов, к-рые должны быть трансдуцированы. Образующиеся трансдуцирующие частицы, по-видимому, вообще не содержат фаговой ДНК.
Для неспецифической Т. характерны следующие особенности: 1) транс-дуцироваться может любой бактериальный ген; 2) включение гена в фаговую частицу может происходить как во время литической инфекции, так и после индуцирования лизогенных клеток на стадии вегетативного фага, а не профага; 3) перенесенный фрагмент хромосомы клетки-донора включается в хромосому клетки -реципиента при участии рекомбинационной системы бактерии, захмещая гомологичный сегмент хромосомы клетки-реципиента; 4) большинство трансдуктантов (по крайней мере, в случае фага Р1), полученных при низкой множественности инфекции, не являются лизогенными и не обнаруживают никаких следов «остаточного» фага.
Каждая трансдуцирующая частица несет и передает клетке-реципиенту лишь незначительную часть генома той клетки-донора, в к-рой она образовалась. Частота котрансдук-ции сцепленных генов тем выше, чем ближе эти гены расположены друг к другу. Максимальное удаление двух генов друг от друга, когда еще возможна котрансдукция (как сказано выше, удаленные гены никогда не трансдуцируются совместно), составляет 2% от общей длины генома E. coli. Гены, расположенные в отрезке, длина к-рого составляет 0,5% генома, трансдуцируются совместно с частотой, достигающей 50% и более.
В 1965 г. Икеда (H. Ikeda) и Томидзава (J. Tomizawa) попытались выяснить причину различия между свойствами трансдуктантов, возникающих при специфической и при неспецифической Т. Ими было показано, что трансдуцирующие частицы фага Р1 либо совсем не содержат фаговой ДНК, либо содержат ее в очень небольшом количестве. Это открытие позволило объяснить, почему трансдуктанты, возникающие при единичном заражении нелизогенных реципиентов, нелизогенны сами и почему индуцирование лпзогенных трансдуктантов, содержащих профаг Р1, не приводит к образованию лизатов HFT. Это связано с тем, что трансдуцирующие частицы фага Р1 содержат фрагменты бактериального генома, не включенные в состав генома фага. Поэтому они неспособны вегетативно размножаться в клетке-реципиенте даже в присутствии инфекционного генома фага-помощника ii не наделяют клетку-реципиент иммунитетом к суперинфекции фагом Р1.
В Р1-лизате ок. 0,3% всех фаговых частиц обладают трансдуцирующей активностью. Полагают, что фрагмент ДНК клетки-хозяина, к-рый может быть включен в трансдуцирующую фаговую частицу, состоит приблизительно из 100 тыс. нуклеотидных пар, что составляет примерно 2,5% длины бактериального генома (см. Дезоксирибонуклеиновые кислоты). Если частота встречаемости трансдуцирующих частиц в фаголизате составляет 0,3%, то частота встречаемости частиц, содержащих какой-либо определенный ген клетки-донора, составляет 0,025-0,003, т. е. 7•10-5. Это довольно хорошо согласуется с тем, что эффективность возникновения трансдуктантов Тгр+ (Тгр — обозначение гена, контролирующего синтез аминокислоты триптофана) при заражении ауксотрофов Тгр~ фагом Р1, размноженным на штамме дикого типа Тгр+, составляет примерно 3*10-5 трансдуктантов на одну адсорбированную частицу фага Р1. Т. о., эффективность интеграции трансдуцирующего фрагмента в данном случае составляет ок. 40%.
Более тщательное изучение судьбы транс дуцирующей ДНК после ее вхождения в реципиентные бактерии показало, что только 7 — 15% трансдуцирующей ДНК оказывается ковалентно связанной с ДНК реципиента. ДНК, остающаяся не связанной, не реплицируется (см. Репликация) и, по-видпмому, представляет собой абортивно трансдуциро-ванную ДНК.
История исследования
Процесс перемещения ДНК был установлен американскими исследователями Д. Ледербергом и Н. Циндером в 1952 г. Ученые изучали функции и строение бактерии Salmonella typhimurium и фага Р22. В результате проведенных опытов они пришли к выводу: происходит генетический обмен между микроорганизмами, не способными к синтезу соединений, и прототрофами.
Между встраивающимися генами не наблюдается физического контакта, процесс не идентичен трансформации. Открытие ученых оказалось случайным, один из штаммов был подвергнут состоянию дезорганизации благодаря вмешательству вириона Р22.
В ходе эксперимента в клетке наблюдали вегетативный рост профага. Через некоторое время образовавшиеся частицы фага проникали в ауксотрофный вид и переносили в его клетки гены своего прежнего хозяина.
Трансдукционное картирование
При переносе бактериальной ДНК вирионы размножаются на чувствительных клетках донора. Для мутагенного ДНК-связывающего белка, содержащего фрагмент нуклеопротеида и несущего наследственную информацию, проводят идентификацию донора и хозяина по нескольким признакам. Для установления сцепления используют отборное скрещивание.
После обработки вирионом структурные единицы организма помещают в питательную среду, не содержащую треонина. В питательной смеси не происходит рост гибридной нуклеиновой кислоты, но образуются структурные единицы, получившие часть хромосомы донора с многофакторным наследственным материалом.
На этом же участке может располагаться второй ген, встречаются двойные Iht+Leu+. Их можно обнаружить, поместив колонии трансдуктантов в питательную среду, не содержащую лейцина. Аминокислоту используют как неселективный отборный материал, необходимый для установления котрансдукции.
Трансдукция
Трансдукция — перенос бактериофагом участка ДНК из одной бактериальной клетки в другую, ведущий к изменению наследственных свойств.
|
Появление колоний вторичного роста после лизиса актинофагом. Культура Act. orientalis продуцирует антибиотик ванкомицин. |
Трансдукция происходит довольно редко: из одного и более миллионов фаговых частиц только одна способна осуществлять трансдук-цию.
Трансдукция — передача инфицирующими бактериальную клетку бактериофагами частей бактериальной хромосомы другим бактериям, которые вследствие этого генетически изменяются.
Трансдукция — явление, аналогичное бактериальным трансформациям. Например, при заражении бактериофагом Т2 мутантной формы кишечной палочки, не способной синтезировать тимин, клетки последней приобретают свойство образовывать фермент, ранее в ней отсутствовавший и необходимый для синтеза тимина. Но ведь при заражении бактериальной клетки в нее проникает только фаговая ДНК, а не белок. Значит, только фаговая ДНК влияет на трансдукцию, наделяющую бактериальную клетку указанным свойством. Однако для бактериофага такая трансдукция характерна лишь тогда, когда он до заражения инкубировался с культурой, способной к такому синтезу. Следовательно, этот процесс аналогичен передаче признаков путем трансформации.
Полная трансдукция, при которой происходит замена аллелей, наблюдается значительно реже, чем так называемая абортивная трансдукция, при которой новый генетичеакий материал поступает в клетку, остается там в течение небольшого периода времени и включается только в часть дочерних клеток.
Трансдукцией называют передачу ДНК от клетки-донора клетке-реципиенту при участии бактериофагов. Обычно при этом фаг переносит лишь небольшой фрагмент ДНК хозяина. Различают два вида транс-дукции: неспецифическую ( общую), при которой может быть перенесен любой фрагмент ДНК хозяина, и специфическую, затрагивающую лишь строго определенные фрагменты ДНК.
При трансдукции геном бактерии-реципиента рекомбинирует с участком генома фага, несущим генетическую информацию от бактерии-донора.
|
Появление колоний вторичного роста после лизиса актинофагом. Культура Act. orientalis продуцирует антибиотик ванкомицин. |
При трансдукции фаг играет роль механического переносчика; лизогенизация клетки не обязательна. Один и тот же фаг может переносить разные свойства.
Явление трансдукции заключается в том, что бактериофаги, растворившие подвергшуюся их нападению бактериальную клетку, переносят части хромосом из этой клетки в другие бактериальные клетки, которые вследствие этого оказываются генетически измененными.
Явление трансдукции, по существу, аналогично трансформации. Разница состоит в том, что в первом случае генетический материал переносится фагами, а во втором происходит непосредственное введение структур, содержащих частицы ДНК.
При обычной трансдукции переносятся любые генетические маркеры, но с различной вероятностью. Только самые близкие генетические локусы могут переноситься при трансдукции одновременно.
|
Типы рецепторов и воспринимаемых ими сигналов. |
Осуществляя трансдукцию, рецептор кодирует сигнал частотой нервных импульсов, которая в ЦНС декодируется и используется для генерирования адекватной реакции.
КООРДИНАЦИЯ И РЕГУЛЯЦИЯ У ЖИВОТНЫХ
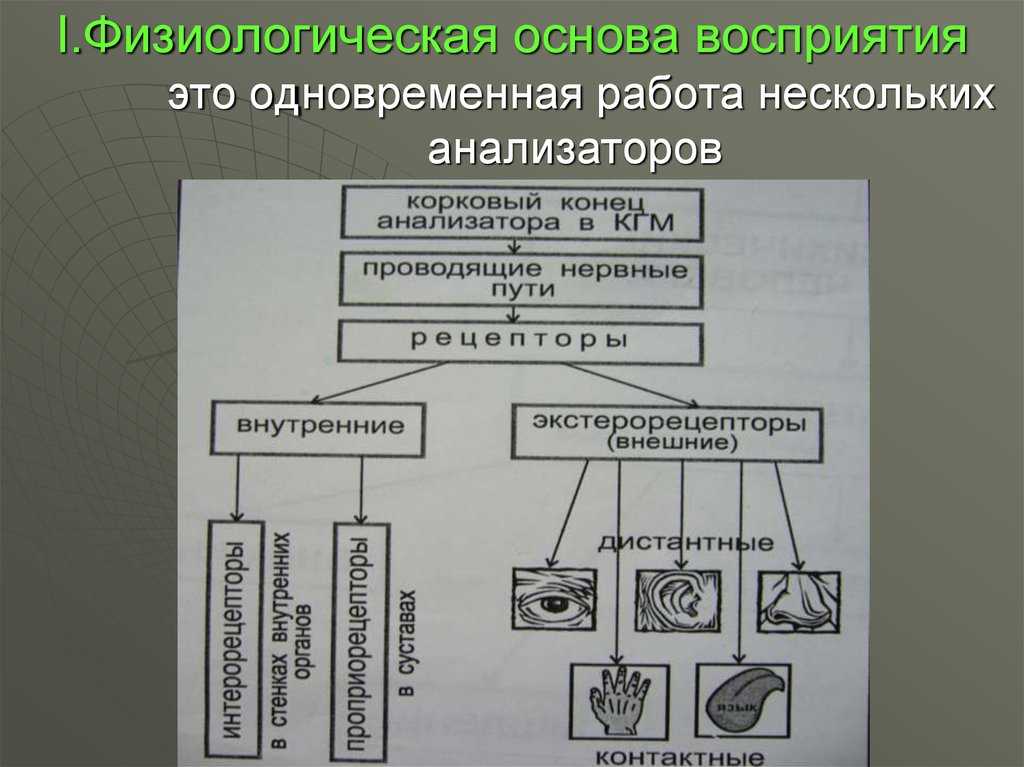
17.4. Сенсорные рецепторы
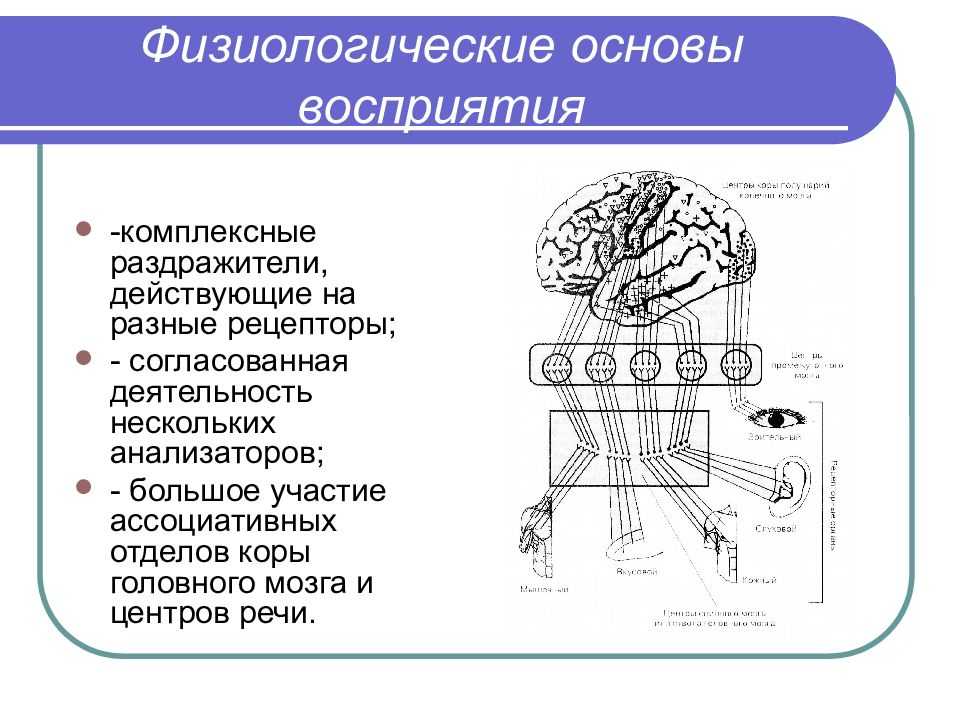
Согласованная работа организма основана на непрерывном поступлении информации от внешней и внутренней среды. Если какая-либо информация вызывает изменение в активности или поведении животного, то ее называют раздражителем или сигналом. Специализированные структуры, воспринимающие раздражители, называются сенсорными рецепторами.
Рецептор простейшего наиболее примитивного типа образован единственным сенсорным нейроном, который способен воспринимать сигналы и порождать нервные импульсы, направляющиеся в мозг. Примером могут служить кожные механорецепторы типа телец Пачини (разд. 17.5.1).




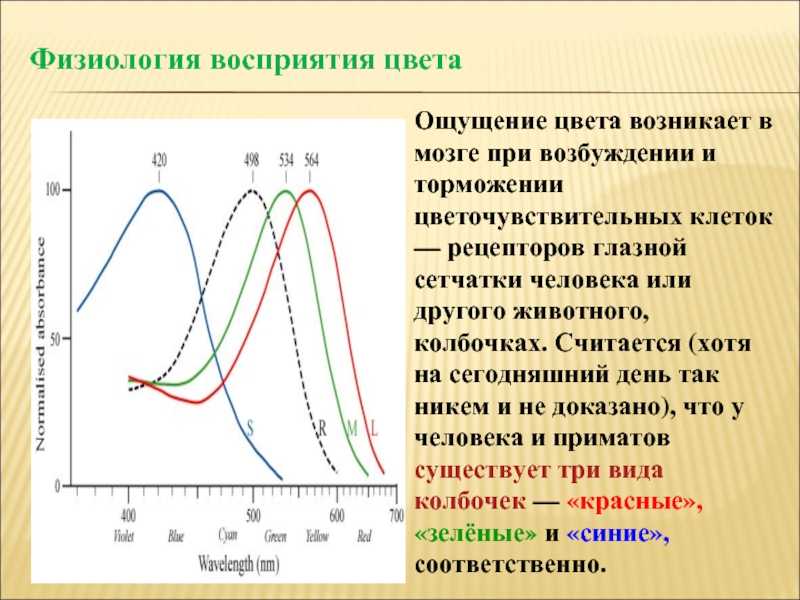
Более сложные — вторичные — рецепторы представлены модифицированными эпителиальными клетками, которые приобрели способность воспринимать сигналы. Эти клетки образуют синапсы с соответствующими сенсорными нейронами, передающими импульсы в ЦНС; таковы, например, палочки и колбочки сетчатки или клетки вкусовых луковиц млекопитающих.
Самые сложные рецепторные структуры — сенсорные органы, или органы чувств, например глаз или ухо. Они состоят из большого числа рецепторных клеток, сенсорных нейронов и связанных с ними вспомогательных структур, служащих для повышения чувствительности и разрешающей способности восприятия. К ним относятся такие структуры, как, например, хрусталик и радужка глаза.
Классификация рецепторов по типам воспринимаемых ими сигналов приведена на табл. 17.6.
Таблица 17.6. Типы рецепторов и воспринимаемых ими сигналов
|
Tип рецептора |
Форма стимулирующей энергии |
Природа сигнала |
|
Фоторецептор |
Электромагнитная |
Свет |
|
Электрорецептор |
Электромагнитная |
Электричество |
|
Механорецептор |
Механическая |
Звук, прикосновение, давление, гравитация, вращение, вибрация |
|
Терморецептор |
Тепловая |
Температура |
|
Хеморецептор |
Химическая |
Влажность, запах, вкус |
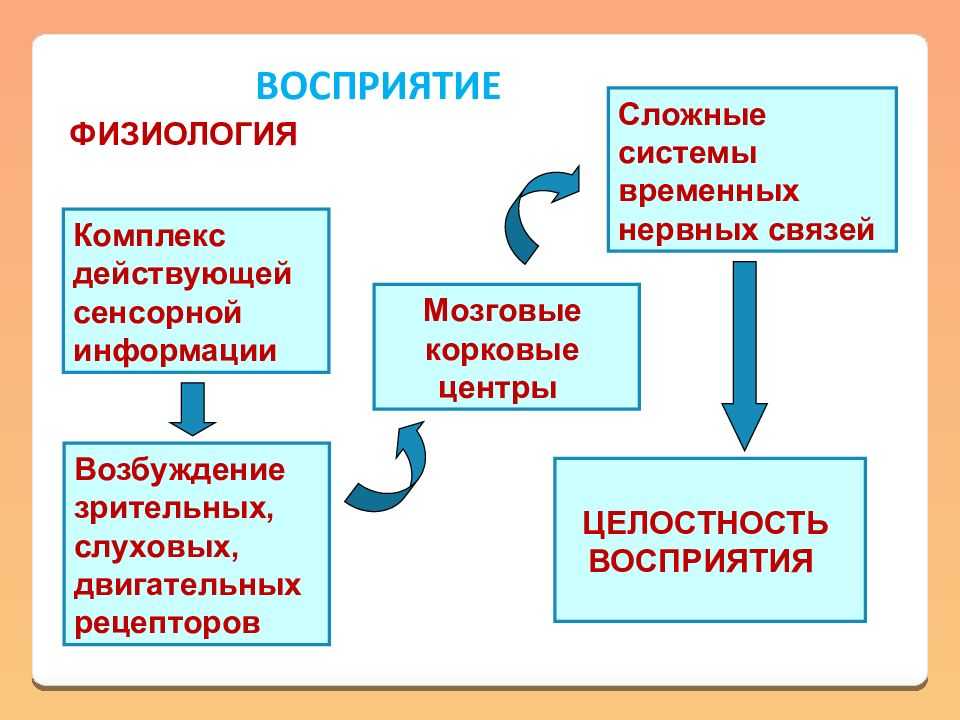
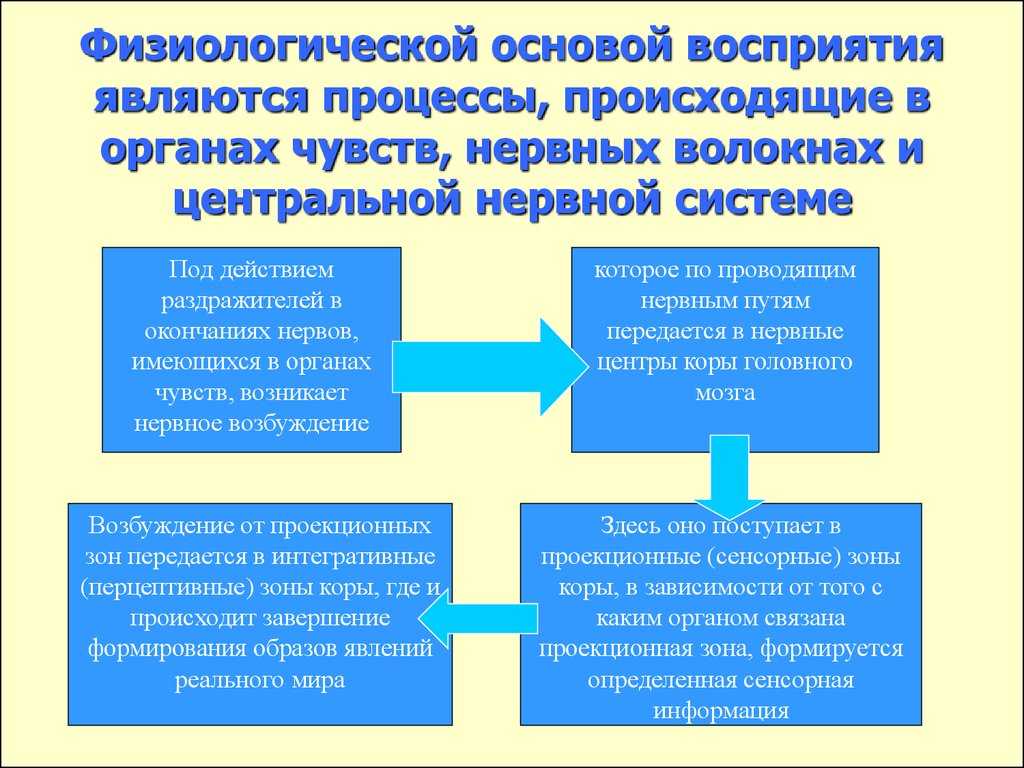
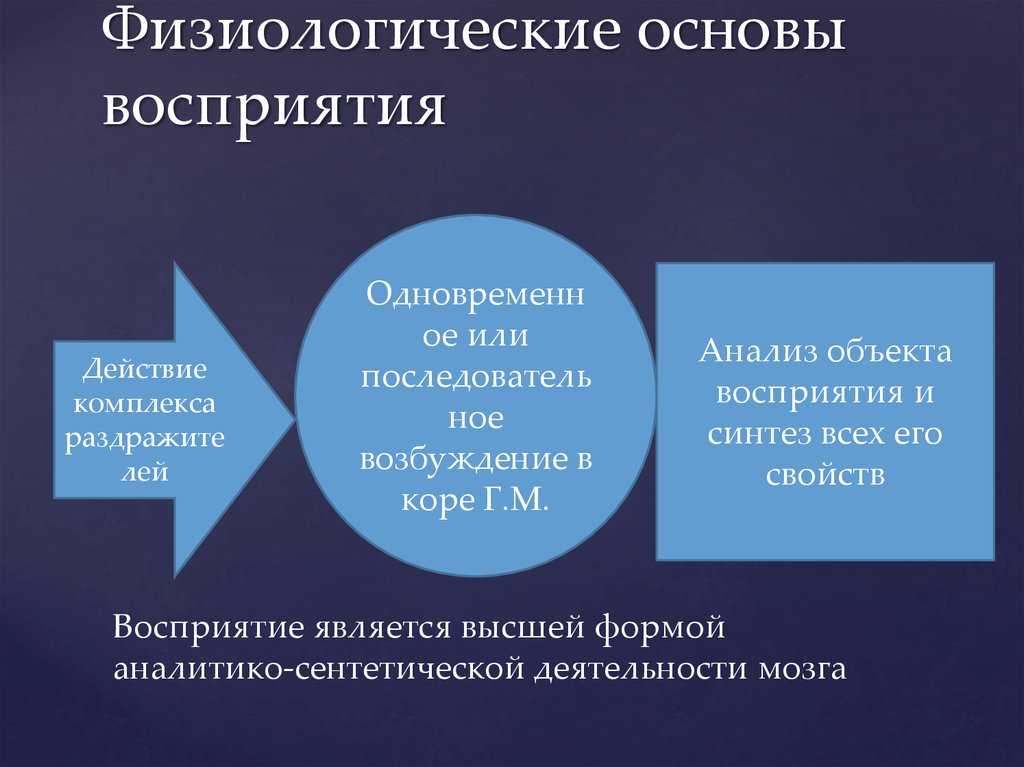
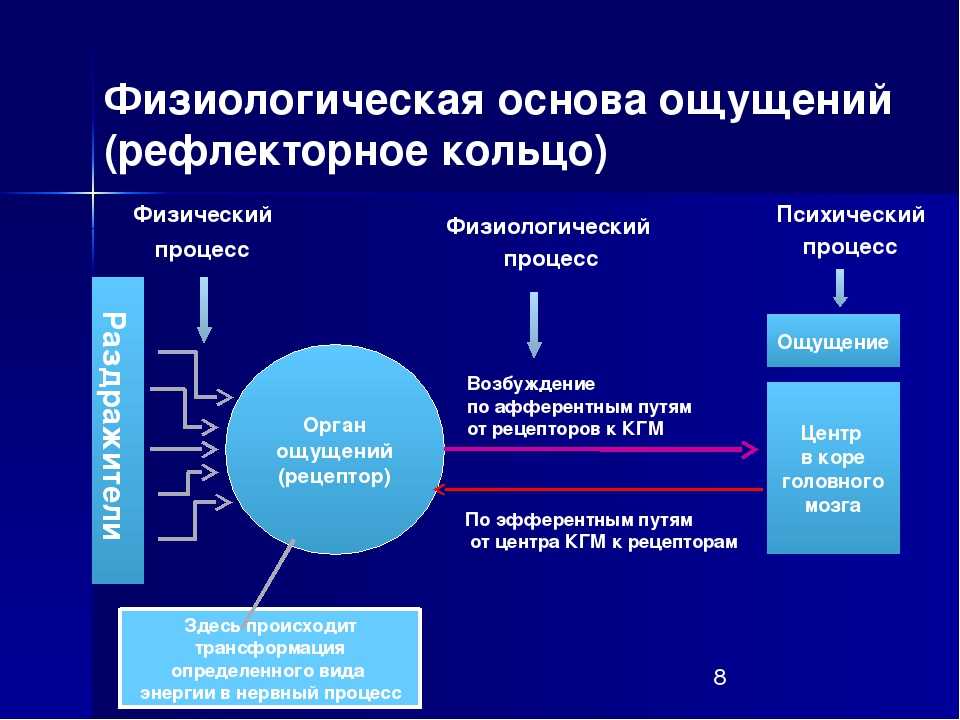
Как следует из этой таблицы, животные чувствительны к разным формам энергии, однако все они должны быть преобразованы в электрическую. Структуры, преобразующие энергию сигнала в электрическую (нервный импульс), распространяющуюся по аксонам, называются биологическими трансдукторами, а сам процесс преобразования — трансдукцией.
Осуществляя трансдукцию, рецептор кодирует сигнал частотой нервных импульсов, которая в ЦНС декодируется и используется для генерирования адекватной реакции.
17.4.1. Механизм трансдукции
Природа воспринимаемых сигналов весьма разнообразна — свет, звук, прикосновение, химическое вещество, но, как уже говорилось, процесс трансдукции превращает их в одинаковые нервные импульсы, электрические по своей природе.
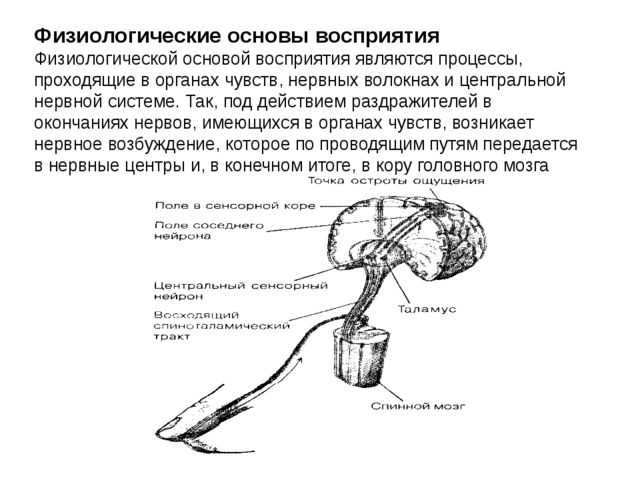
Все рецепторы представляют собой возбудимые клетки, т. е. подобно нейронам и мышечным волокнам они реагируют на соответствующий им сигнал быстрым изменением электрических свойств своей мембраны. В отсутствие стимуляции они сохраняют потенциал покоя, описанный в разд. 17.1.1. Сигнал вызывает изменение мембранного потенциала. Бернард Кац в 1950 г. при изучении сложного рецептора растяжения — мышечного веретена — продемонстрировал его деполяризацию в области, прилегающей к окончаниям сенсорных нейронов. Такая местная деполяризация обнаруживается только в рецепторной клетке и называется генераторным потенциалом. Дальнейшие исследования с использованием микроэлектродов, введенных в рецепторные клетки мышечных веретен и кожные механорецепторы (тельца Пачини) позволили получить следующие данные о трансдукции:


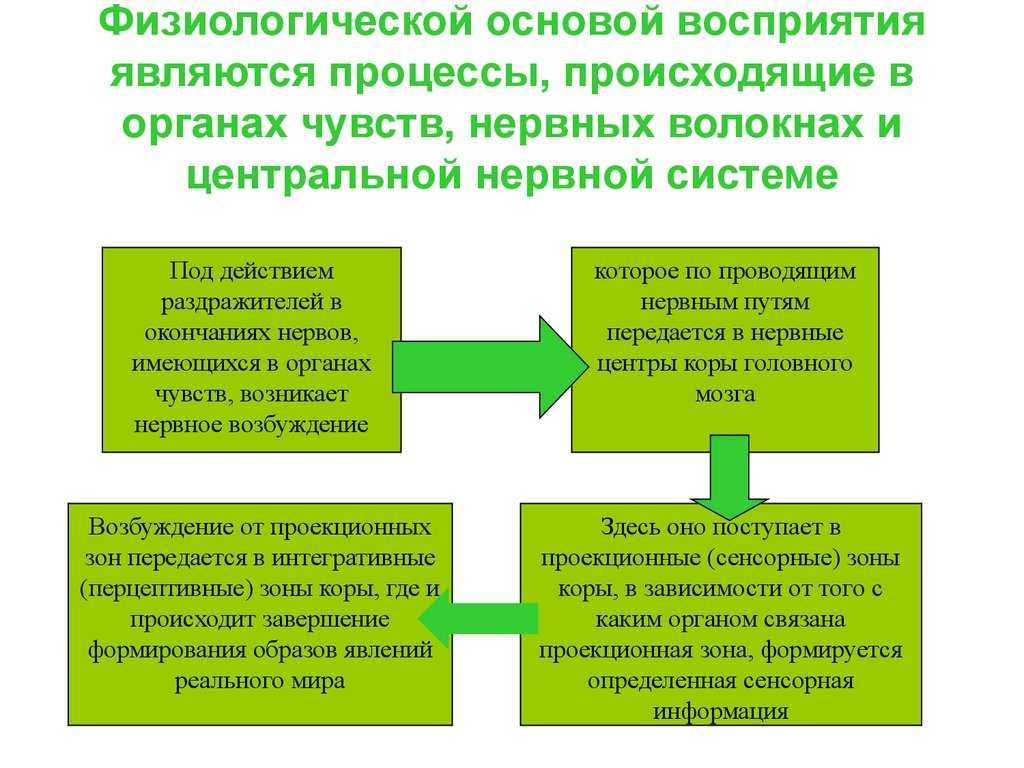




1) генераторный потенциал возникает при действии сигнала, повышающего проницаемость мембраны рецепторной клетки для ионов натрия и калия, которые в результате движутся по своим электрохимическим градиентам;
2) величина генераторного потенциала зависит от интенсивности сигнала;
3) когда генераторный потенциал достигает порогового уровня, он генерирует потенциал действия (рис. 17.30);
4) частота импульсов в сенсорном аксоне прямо пропорциональна интенсивности сигнала.
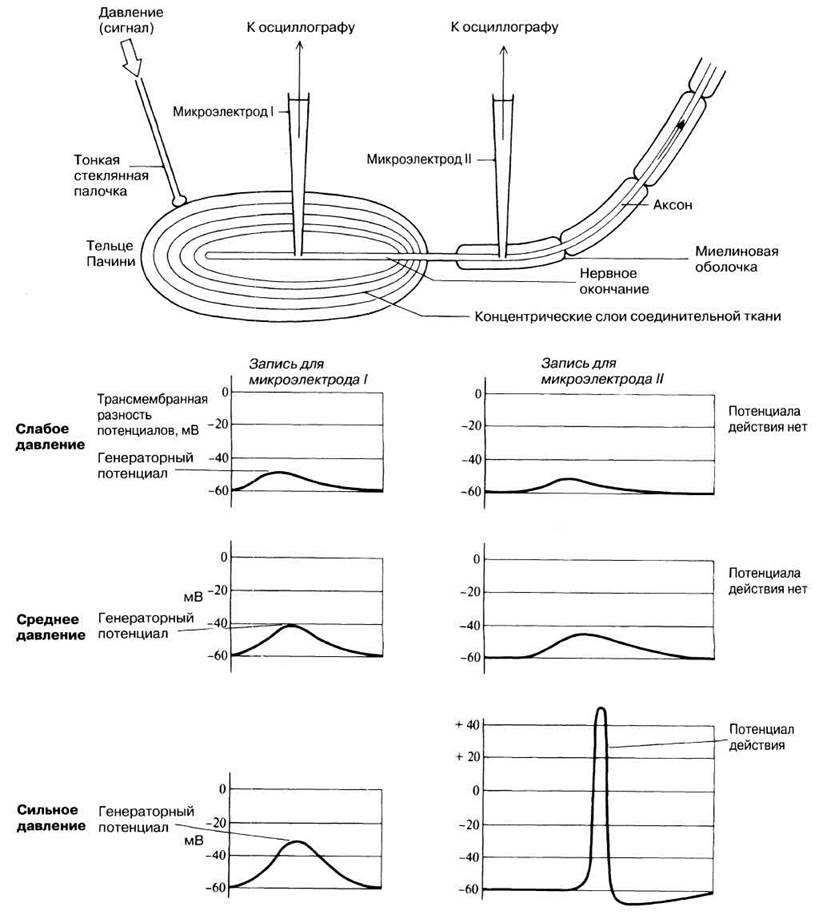
Рис. 17.30. Электрическая активность, зарегистрированная двумя микроэлектродами, один из которых (I) введен в окончание аксона сенсорного нейрона, находящегося внутри тельца Пачини, а второй (II) — в тот же аксон, но уже за пределами тельца Пачини. По мере того как надавливание тонкой стеклянной палочкой (сигнал) увеличивается, растет и генераторный потенциал, который, достигнув порогового уровня, возбуждает в сенсорном нейроне потенциал действия.
ПредыдущаяСледующая
Вирусы звучащего
Перевод энергии звучащего в иные формы (семиотические или материальные) предполагает открытие вибрационных потенциалов и их кодирование в новых агрегатных состояниях. Вечная пифагорейская мечта человечества о дешифровке бесконечности космической музыки, перенесенная в индустрию электронного воспроизводства и цифрового анализа параметров звуковых объектов, — это не просто жажда присвоения (или использования) звучащего, но и постоянное рассечение и выявление в нем каналов восприятия, распространения, инструментализации и машинизации функций для картографирования того, что ранее не рассматривалось как часть знания. «Радио существовало до того, как было изобретено, а до того, как оно было услышано, оно было», — по утверждению Дугласа Кана, опыты прослушивания Т. Ватсоном, ассистентом А.Г. Белла, шумов телефонных линий были встречей ученого с электромагнитными волнами . Именно это открытие позволило продвинуться в установлении логики совокупности процессов, оборачивающих планетарный масштаб экологии — за десять лет до уточнения их реальности Г. Герцем и задолго до получения первых снимков Земли из космоса. В этом событии ситуативность проявления звучащего неразрывно связана с его процессуальностью.
Потенциальные силы звучащего способны проникать, как вирусные механизмы, в структуры знания. Вирусы звучащего, таким образом, артикулируют себя в промежутке между знанием и мышлением, составляя и с тем, и с другим новые эпистемические модели.
Трансдукция конструирует цепи связности материальных событий с их эффектами через картографию переходов и скачков между уровнями выразительности. Этот процесс, обеспечивающий напряжение и дистанцию между содержанием и выражением, заражает установленные абстрактные машины звучащего новыми техниками. Вирусы звучащего усложняют формирование территорий трансдукции: появление акустического тела или органа, смещение ритма, собирающее новые звуковые объекты, — все эти неизведанные области уже реализуют себя в ситуации натурализованных абстрактных машин схватывания и воспроизводства звуковых материалов, но требуют постоянной пересборки, распространения и конструирования аномальных ситуаций в экологии заражения. Это движение искажает логику кодировки, обеспечивая мышление композиционной экспериментальностью действия. Вопрос о технике звучащего, таким образом, становится проблемой выявления автономных концептов, способных не просто делать звучащее более узнаваемым, но и вскрывать в нем пробелы, слепые пятна и лакуны, удерживающие его от кристаллизации.
B. Специализированные трансдукционные переносы – определенные донорские гены
- При специализированной трансдукции трансдуцирующие частицы несут только определенные части бактериального генома. Дефект в лизогенном жизненном цикле фагов умеренного пояса, которые встраивают свои геномы в определенный участок хромосомы хозяина, делает возможной специализированную трансдукцию.
- Когда профаг вынужден покинуть свою хромосому-хозяин, вырезание иногда выполняется неправильно. Как и в случае с F’-плазмидами, полученный фаговый геном содержит от 5 до 10 процентов бактериальной хромосомы, примыкающей к месту интеграции.
- Однако трансдуцирующая частица дефектна, потому что в ней отсутствуют вирусные гены, и поэтому она не может воспроизводиться независимо. Несмотря на это, он вводит оставшийся вирусный геном, а также любые бактериальные гены, которые он переносит, в другую бактерию.
- При правильных условиях бактериальные геномы могут стать постоянно интегрированными. Фаг E. coli lambda является наиболее тщательно изученным примером специализированной трансдукции.
- В определенных местах, известных как сайты прикрепления или att, геном лямбда интегрируется в хромосому хозяина. Сайты att фагов и сайты att бактерий сопоставимы и могут образовывать комплексы.
- Сайт att для лямбда расположен между генами gal и bio на хромосоме E. coli; следовательно, эти бактериальные гены часто присутствуют, когда лямбда вырезается неправильно для создания специализированной трансдуцирующей частицы.
- Продукт лизиса клеток (лизат), полученный в результате индукции популяции лизогенизированных клеток E. coli, содержит нормальный фаг и несколько дефектных трансдуцирующих частиц.
Специализированная трансдукция умеренным бактериофагом. | Кредит изображения: Микробиология Прескотта Джоанны Уилли (автор), Линды Шервуд (автор), Кристофера Дж. Вулвертона (автор)
Пример специализированной трансдукции
- Бактерия фага E. coli лямбда является наиболее хорошо изученным примером специализированной трансдукции.
- Ген лямбда встраивается в хромосому хозяина в определенных местах, называемых точками прикрепления или точками присоединения.
- Бактериальные и фаговые сайты очень похожи и могут взаимодействовать друг с другом.
- Сайт att лямбда находится между генами bio и gal на хромосоме E.coli. Это означает, что когда лямбда вырезается неправильно, чаще всего присутствует эта бактериальная генетика.
- Продукт лизиса клеток (лизат), полученный в результате индукции ряда лизогенизированных клеток бактерий E. coli, содержит как нормальный фаг, так и несколько дефектных трансдуцирующих частиц.
- Эти частицы называются лямбда-гал, если они обладают генами использования галактозы, или лямбда-би, если они имеют био с противоположной стороны атта.
Механизм трансдукции фага лямбда и кишечной палочки.
Между генами gal и bio интегрирован лямбда-фаг. Если он возбуждается нормально (вверху слева), то новый фаг не содержит бактериального гена и является полным. В редких случаях вырезание выполняется асимметрично (вверху в центре), что означает, что либо ген gal, либо биогены поглощаются, и часть генетики фага теряется. Гены gal являются единственным исключением из этого правила. Создается дефектный лямбдафаг, который несет бактериальную генетику и может передать ее новому реципиенту.
Механизм трансдукции фага лямбда и кишечной палочки. — схема бактериальной трансдукции
Этапы специализированной трансдукции
- Во время лизогенного цикла ДНК фага интегрируется в бактериальную хромосому после инфицирования бактериофагом донорской бактерии.
- В результате неточного разделения ДНК фага также удаляется часть бактериальной хромосомы.
- Затем фаг, содержащий часть бактериальной хромосомы, заражает нового хозяина, и во время лизогенного цикла репликации ДНК донора включается в бактерию-реципиент.
- Затем реципиент демонстрирует недавно приобретенную генетическую характеристику.
Этапы специализированной трансдукции
FAQ
Что такое бактериальная трансдукция?
Трансдукция — это процесс, с помощью которого вирус передает генетический материал от одной бактерии к другой. Вирусы, называемые бактериофагами, способны инфицировать бактериальные клетки и использовать их в качестве хозяев для создания большего количества вирусов.
Чем трансдукция отличается от других способов генетического переноса у бактерий?
Трансдукция отличается от других способов генетического переноса, таких как конъюгация и трансформация, поскольку она включает перенос ДНК, опосредованный бактериофагами.
Какие существуют виды трансдукции?
Существует два типа трансдукции: генерализованная трансдукция и специализированная трансдукция. Генерализованная трансдукция включает случайный перенос любой бактериальной ДНК, тогда как специализированная трансдукция включает перенос определенных последовательностей ДНК, прилегающих к интегрированному профагу.
Какова роль бактериофагов в трансдукции?
Бактериофаги служат транспортными средствами для переноса бактериальной ДНК во время трансдукции. Они заражают бактериальные клетки, приобретают фрагменты бактериальной ДНК и затем доставляют эту ДНК бактериям-реципиентам.
Как происходит генерализованная трансдукция?
При генерализованной трансдукции во время литического цикла репликации бактериофага случайные фрагменты бактериальной ДНК ошибочно упаковываются во вновь образованные фаговые частицы. Затем эти фаговые частицы заражают другие бактерии, перенося бактериальную ДНК.
Как происходит специализированная трансдукция?
Специализированная трансдукция происходит, когда профаг (интегрированный фаговый геном) вырезается из бактериальной хромосомы, но несет с собой соседнюю бактериальную ДНК. Этот вырезанный профаг, теперь содержащий бактериальную ДНК, заражает другие бактерии и переносит специфические последовательности ДНК.
Каково значение трансдукции в эволюции бактерий?
Трансдукция играет решающую роль в эволюции бактерий, способствуя передаче генетического материала, включая полезные признаки и гены устойчивости к антибиотикам, между бактериальными штаммами или видами.
Может ли трансдукция передавать определенные гены или генетические маркеры?
Да, трансдукция может передавать определенные гены или генетические маркеры в зависимости от типа трансдукции. Специализированная трансдукция может передавать определенные последовательности ДНК, соседние с интегрированным профагом, в то время как генерализованная трансдукция может передавать случайные фрагменты ДНК.
Чем трансдукция отличается от горизонтального переноса генов?
Трансдукция — это один из механизмов горизонтального переноса генов, который относится к передаче генетического материала между организмами одного поколения. Трансдукция, в частности, включает перенос бактериальной ДНК через бактериофаги.
Каковы применения трансдукции в исследованиях и биотехнологии?
Трансдукция имеет различные применения в генетических исследованиях, включая генетическое картирование, перенос генов и изучение устойчивости к антибиотикам. Он также используется в биотехнологии для генной инженерии, генной терапии и создания трансгенных организмов.
Что такое трансдукция?
Давайте начнем с некоторых основных словарных определений.
Преобразовать означает преобразовать что-то в другую форму.
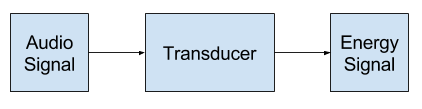
Это популярный термин из области электроники и обработки сигналов, где «преобразователь”- это общее название для компонентов или модулей, преобразующих звуки в энергию или наоборот.
Итак, как правило, мы видим, что преобразование — это преобразование сигнала в другую форму.
Описание обработки сигнала является наиболее заметным, когда звуковые волны превращаются в электрическую энергию для некоторого использования в системе. Каждый звук будет представлен некоторой электрической сигнатурой на некотором выбранном уровне дискретизации.







Рекомендации
- https://en.wikipedia.org/wiki/Transduction_(genetics)
- https://www.nature.com/scitable/definition/transduction-prokaryotes-292/
- https://www.britannica.com/science/transduction-microbiology
- https://study.com/academy/lesson/bacterial-transduction-definition-process-advantages.html
- https://www.khanacademy.org/science/ap-biology/gene-expression-and-regulation/mutations-ap/a/genetic-variation-in-prokaryotes
- https://www.slideshare.net/8988337117/vikas-pathania
- https://www.vedantu.com/biology/transduction-microbiology
- https://www.bioexplorer.net/bacterial-transduction.html/
- https://blog.addgene.org/plasmids-101-transformation-transduction-bacterial-conjugation-and-transfection
Специфическая трансдукция
Ученые изучили перенос бактериальной ДНК вирусом Л. Фаг располагается на att-участке нуклеопротеидной структуры Е.coli, имеющей одинаковую последовательность фосфорных эфиров.
В период выведения вириона его исчезновение часто сопровождается ошибкой: удаляется часть, по размеру равная фрагменту ДНК. Особенность процесса состоит в том, что начало обломка располагается в другом месте. Некоторые гены вируса исчезают, другие наследственные факторы, несущие информацию, поглощаются фагом.
Некоторые фаги внедряются в любую часть нуклеопротеиновой структуры и переносят наследственную информацию, используя механизм специфического перемещения бактериальной ДНК из одной клетки в другую.
В хромосоме находится пространство, имеющее общие черты с att-участком и ДНК вируса. При разрушении гомологичного ресурса осуществляют внедрения фага в нуклеопротеид и его передачу при специфическом перемещении генов, расположенных рядом.
Фаги переносят наследственную информацию.
Формирование частиц фага лямбда
Вирус принадлежит к умеренным фагам E.coli, обладающим двухцепочной ДНК. Фаг внедряется в генную структуру бактерий в виде профага и передается нескольким поколениям.
Вирион лямбда состоит из следующих элементов: головки, хвоста, боковых нитей, белковой пленки. Внедрение сопровождается генетическим перераспределением материала на участке бактерии. В результате обмена наследственной информацией в присутствии вирусного и бактериального белка формируется форма Холлидея.
Вирус лямбда имеет 2 пути развития — мутагенный и литический. Фаг образует новые частицы, которые попадают во внешнюю среду, а клетка хозяина гибнет. В случае развития лизогенной реакции геном вириона проникает в хромосому E.coli и передается нескольким поколениям.
Какие события происходят в клетке реципиента
Вирус Л перемещает хромосомы от клеток донора к реципиенту. Ученые вывели геном фага, внедренного в хромосомную ДНК бактерии в культуре клеток кишечной бактерии К 12. В процессе получили расщепленные вирусом структурные элементы. Ими заражали культуры типа К 12. Затем их высевали в питательные среды и учитывали количество колоний, проверяя наличие аллелей от клеток-доноров, передавшихся мутантным реципиентам.
Ученые вывели геном фага, внедренного в хромосомную ДНК.
Вирион может переносить часть хромосомы, но процесс специфический, его осуществляет геном gal, находящийся на хромосоме бактериальной структуры. Многие трансдуктанты Cal+ нестабильны, теряют этот признак.
Различия между трансдукцией и передачей
| трансдукция | коробка передач | |
|---|---|---|
| Определение | Процесс переноса ДНК, опосредованный бактериофагами | Процесс распространения болезни или патогена от одного хозяина к другому |
| Биологический контекст | Встречается у бактерий | Встречается у различных организмов (например, у животных, растений, человека) |
| Генетический материал | Перенос фрагментов бактериальной ДНК | Перенос ДНК возбудителя или целых инфекционных агентов |
| Механизм | Опосредуется бактериофагами | Опосредуется различными путями (например, прямой контакт, воздушно-капельный, переносчики) |
| Трансферный маршрут | Внутри бактериальных популяций | Между хостами или отдельными лицами |
| Диапазон хостов | Бактериальные клетки | Различные микроорганизмы в зависимости от конкретного заболевания или возбудителя |
| природа | Может происходить естественным путем или в лабораторных условиях | Естественное возникновение при инфекционных заболеваниях |
| Примеры | Трансдукция в бактериальных популяциях | Передача вирусов, бактерий, паразитов и др. при инфекционных заболеваниях |
| Факторы, влияющие | Взаимодействие бактериального фага с хозяином, частота инфицирования | Способ передачи, факторы окружающей среды, восприимчивость хозяина |
| Контрольные меры | Понимание и управление взаимодействием фагов в бактериальных популяциях | Вмешательства в области общественного здравоохранения, вакцинация, борьба с переносчиками болезней, соблюдение правил гигиены |