
![Эволюция растений: огромная проблема для приверженцев эволюции [часть i] - origins.org.ua](http://lesniepolyani.ru/wp-content/uploads/3/7/3/37340e16b2bb3de5c33582bc35318a8a.jpeg)
ссылки
- Баум Д.А., Смит С.Д. и Донован С.С. (2005). Древовидный вызов. наука, 310(5750), 979-980.
- Curtis, H. & Barnes, N.S. (1994). Приглашение к биологии. Macmillan.
- Холл Б.К. (ред.). (2012). Гомология: иерархическая основа сравнительной биологии. Академическая пресса.
- Hickman, C.P., Roberts, L.S., Larson, A., Ober, W.C., & Garrison, C. (2001). Интегрированные принципы зоологии. McGraw-Hill.
- Hinchliff, CE, Smith, SA, Allman, JF, Burleigh, JG, Chaudhary, R., Coghill, LM, Crandall, KA, Deng, J., Drew, BT, Gazis, R., Gude, K., Hibbett, Д.С., Кац Л.А., Лафингхаус, Х.Д., Мактавиш, Э.Дж., Мидфорд, П.Е., Оуэн, К.Л., Ри, Р.Х., Рис, Дж.А., Солтис, Д.Е., Уильямс, Т., … Крэнстон, К.А. Синтез филогении и таксономии во всеобъемлющее древо жизни. Слушания Национальной Академии Наук Соединенных Штатов Америки, 112(41), 12764-9.
- Кардонг, К. В. (2006). Позвоночные: сравнительная анатомия, функции, эволюция. McGraw-Hill.
- Пейдж Р.Д. и Холмс Э.С. (2009). Молекулярная эволюция: филогенетический подход. Джон Вили и сыновья.
Ссылки [ править ]
- ^ Летунич, Ивица; Борк, Пер (1 января 2007 г.). «Интерактивное древо жизни (iTOL): онлайн-инструмент для отображения и аннотации филогенетического дерева» . Биоинформатика . 23 (1): 127–128. DOI : 10.1093 / биоинформатики / btl529 . ISSN 1367-4803 . PMID 17050570 . Архивировано 29 ноября 2015 года . Проверено 21 июля 2015 .
- ^ Ciccarelli, FD; Doerks, T .; Фон Меринг, С .; Криви, CJ; Snel, B .; Борк, П. (2006). «К автоматической реконструкции дерева жизни с высоким разрешением» . Наука . 311 (5765): 1283–1287. Bibcode2006Sci … 311.1283C . CiteSeerX 10.1.1.381.95 14 . DOI10.1126 / science.1123061 . PMID 16513982 . S2CID 1615592 .
- ^ а б в г Фельзенштейн Дж. (2004). Вывод о филогенезе Sinauer Associates: Сандерленд, Массачусетс.
- ^ Байи, Анатоль (1981-01-01). Abrégé du dictionnaire grec français . Париж: Ашетт. ISBN 978-2010035289. OCLC 461974285 .
- ↑ Байи, Анатоль. «Греко-французский словарь онлайн» . www.tabularium.be . Архивировано 21 апреля 2014 года . Проверено 2 марта 2018 года .
- Перейти ↑ Hodge T, Cope M (1 октября 2000 г.). «Генеалогическое древо миозинов» . J Cell Sci . 113 (19): 3353–4. PMID 10984423 . Архивировано 30 сентября 2007 года.
- ^ Дерево «Факты: Внедренный против некорневых деревьев» . Архивировано 14 апреля 2014 года . Проверено 26 мая 2014 .
- Перейти ↑ Maher BA (2002). «Выкорчевывание древа жизни» . Ученый . 16 (2): 90–95. Bibcode2000SciAm.282b..90D . DOI10.1038 / Scientificamerican0200-90 . PMID 10710791 . Архивировано 2 октября 2003 года.
- ^ ван Овен, Маннис; Кайзер, Манфред (2009). «Обновленное комплексное филогенетическое дерево глобальных вариаций митохондриальной ДНК человека» . Мутация человека . 30 (2): E386 – E394. DOI10.1002 / humu.20921 . PMID 18853457 . S2CID 27566749 .
- ^ a b c Фельзенштейн, Джозеф (1978-03-01). «Число эволюционных деревьев» . Систематическая биология . 27 (1): 27–33. DOI10.2307 / 2412810 . ISSN 1063-5157 . JSTOR 2412810 .
- ^ Фокс, Эмили. «Дендрограмма» . coursea . конечно Архивировано 28 сентября 2017 года . Проверено 28 сентября 2017 года .
- ^ Майр, Эрнст (2009) «Кладистический анализ или кладистическая классификация?». Журнал зоологической систематики и эволюционных исследований. 12: 94–128. DOI: 10.1111 / j.1439-0469.1974.tb00160.x ..
- ^ Соарес, Антонио; Рабело, Рикардо; Дельбем, Александр (2017). «Оптимизация на основе анализа филограмм». Экспертные системы с приложениями . 78 : 32–50. DOI10.1016 / j.eswa.2017.02.012 . ISSN 0957-4174 .
- ^ Лабандейра, CC; Дилчер, Д.Л .; Дэвис, Д.Р .; Вагнер, DL (1994-12-06). «Девяносто семь миллионов лет ассоциации покрытосеменных и насекомых: палеобиологическое понимание значения коэволюции» . Труды Национальной академии наук . 91 (25): 12278–12282. Bibcode1994PNAS … 9112278L . DOI10.1073 / pnas.91.25.12278 . ISSN 0027-8424 . PMC 45420 . PMID 11607501 .
- ^ Santamaria, R .; Терон, Р. (26 мая 2009 г.). «Treevolution: визуальный анализ филогенетических деревьев» . Биоинформатика . 25 (15): 1970–1971. DOI10.1093 / биоинформатики / btp333 . PMID 19470585 .
- ^ a b «Эволюционная систематика: схемы шпинделя» . Palaeos.com . 2014-11-10 . Проверено 7 ноября 2019 .
- ^ «Деревья, пузыри и копыта» . Мозг трехфунтовой обезьяны — биология, программирование, лингвистика, филогения, систематика… . 2007-11-21 . Проверено 7 ноября 2019 .
- ^ a b c Подани, Янош (2019-06-01). «Коралл жизни» . Эволюционная биология . 46 (2): 123–144. DOI10.1007 / s11692-019-09474-ш . ISSN 1934-2845 .
- ^ Дарвин, Чарльз (1837). Notebook B . п. 25.
- ^ Пенни, D .; Хенди, доктор медицины; Сталь, Массачусетс (1992). «Прогресс в методах построения эволюционных деревьев». Тенденции в экологии и эволюции . 7 (3): 73–79. DOI10.1016 / 0169-5347 (92) 90244-6 . PMID 21235960 .
- ↑ A. Dress, KT Huber и V. Moulton. 2001. Метрические пространства в чистой и прикладной математике. Documenta Mathematica LSU 2001 : 121-139
- ^ Таунсенд ДП, Су Z, ТЕКЛЕ Y (2012). «Филогенетический сигнал и шум: предсказание силы набора данных для разрешения филогении» . Генетика . 61 (5): 835–849. DOI10.1093 / sysbio / sys036 . PMID 22389443 .
- Перейти ↑ Arenas M, Posada D (2010). «Влияние рекомбинации на реконструкцию наследственных последовательностей» . Генетика . 184 (4): 1133–1139. DOI10.1534 / genetics.109.113423 . PMC 2865913 . PMID 20124027 .
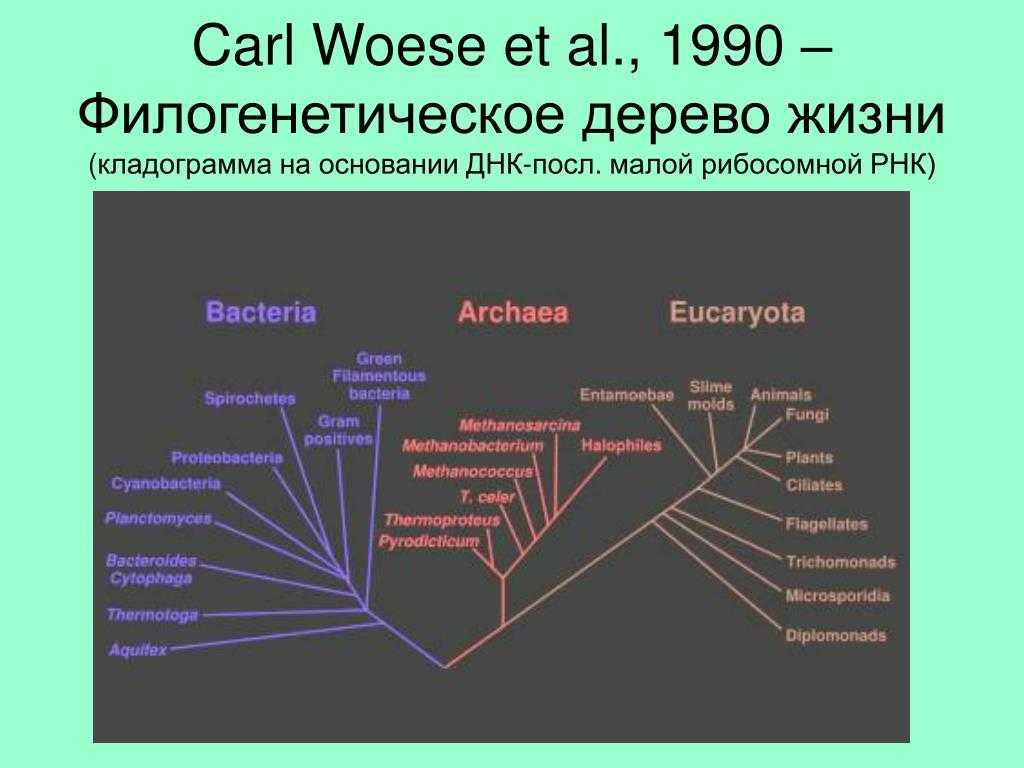
- ^ Вёзе C (2002). «Об эволюции клеток» . Proc Natl Acad Sci USA . 99 (13): 8742–7. Bibcode2002PNAS … 99.8742W . DOI10.1073 / pnas.132266999 . PMC 124369 . PMID 12077305 .
- ^ Parhi, J .; Трипатия, PS; Priyadarshi, H .; Мандал, SC; Панди, PK (2019). «Диагностика митогенома для устойчивой филогении: случай группы рыб Cypriniformes». Джин . 713 : 143967. DOI10.1016 / j.gene.2019.143967 . PMID 31279710 .
Презентация на тему: » Тема: Молекулярные доказательства эволюции Задачи: Сформировать знания о фактах, свидетельствующих в пользу эволюции. Дать краткую характеристику молекулярных,» — Транскрипт:
1
Тема: Молекулярные доказательства эволюции Задачи: Сформировать знания о фактах, свидетельствующих в пользу эволюции. Дать краткую характеристику молекулярных, эмбриологических, сравнительно-анатомических, палеонтологических и биогеографических доказательств эволюции
2
Универсальность генетического кода свидетельствует о едином происхождении всех живых организмов Земли. Данные молекулярной биологии
3
Чем ближе родство, тем меньше отличий в строении ДНК и белков. В настоящее время проведена гибридизация цепей нуклеотидов ДНК человека и шимпанзе. Для этого были разделены двойные цепи ДНК человека и шимпанзе, и затем одиночные цепи ДНК человека соединили с цепями нуклеотидов шимпанзе. Между комплементарными нуклеотидами восстановились химические связи, и оказалось, что ДНК человека и шимпанзе сходны на 91 97,5%. Данные молекулярной биологии
4
В таблице приведены последовательности нуклеотидов в генах митохондриальной ДНК человека, орангутана и ревуна. Постройте их филогенетическое древо, исходя из предположения, что чем больше нуклеотидов пару видов, тем раньше они дивергировали. Реконструируйте последовательность нуклеотидов в гене последнего общего предка этих видов. Данные молекулярной биологии ОрангутанАТГТТЦ-ГЦЦГАЦ-ЦГЦТГГ-ЦТАТТЦ-ТЦЦАЦГ-ААЦЦАЦ-АААГАТ- АТТГГА РевунАТГТТЦ-АТАААТ-ЦГЦТГА-ЦТАТТТ-ТЦААЦТ-ААЦЦАТ-АААГАТ- АТТГГТ ЧеловекАТГТТЦ-ГЦЦГАЦ-ЦГТТГА-ЦТАТТЦ-ТЦТАЦА-ААЦЦАЦ-АААГАЦ-АТТГГА Строим матрицу различий ЧеловекОрангутан Ревун
5
На основе матрицы различий строим филогенетическое древо. Рассуждая, между человеком и орангутаном мы обнаружили меньше рразличий, чем между ревуном и человеком, и ревуном и орангутаном.. Следовательно, эволюционные линии человека и орангутана разделились позже, чем линии ревуна и общего предка человека и орангутана. Узлы ветвления обозначают точку дивергенции, т.е. время, когда на Земле жил последний общий предок двух или более разных эволюционных линий. Длина ветвей пропорциональна числу накопленных изменений
В филогенетических древах важно положение точек ветвления и длина ветвей, но не направление ветвей. Данные молекулярной биологии. 6
6
Предковую последовательность восстанавливаем по позициям, сходным у большинства видов Данные молекулярной биологии ПредокАТГТТЦ-ГЦЦГАЦ-ЦГЦТГА-ЦТАТТЦ-ТЦ?АЦ?-ААЦЦАЦ-АААГАТ-АТТГГА Домашнее задание АкулаЦГЦТЦТ-ТАГТЦЦ-ЦАЦГТГ-ГГАЦЦЦ-ТЦАЦЦТ-ТАГЦТА-АГАТЦА-ЦТАЦЦГ-ТА МышьЦЦЦТЦТ-ТАЦТЦЦ-ЦАЦГТГ-ГГАЦЦЦ-ТЦТЦЦТ-ТАГЦТА-ТГАТЦА-ЦААЦЦЦ-ТА ОпоссумЦЦЦТЦТ-ТАЦТГГ-ГТГЦАЦ-ЦЦАЦЦЦ-ТЦТЦЦТ-ТАГЦТА-ТГАТЦА-ЦТАЦЦЦ-ТА
7
Древо 1 построено на основе сравнения последовательностей ДНК. Какое из древ 2-5 изображено неверно? Ответ: 5
История
Идея « древа жизни » возникла из древних представлений о лестничном переходе от низших форм жизни к высшим (например, в Великой Цепи Бытия ). Ранние изображения «ветвящихся» филогенетических деревьев включают «палеонтологическую карту», показывающую геологические взаимоотношения между растениями и животными в книге Эдварда Хичкока « Элементарная геология » (первое издание: 1840 г.).
Чарльз Дарвин (1859) также создал одну из первых иллюстраций и чрезвычайно популяризировал понятие эволюционного «дерева» в своей основополагающей книге «Происхождение видов» . Более века спустя, эволюционные биологи до сих пор используют диаграммы дерева для описания эволюции , поскольку такие схемы эффективно передать концепцию , что видообразование происходит через адаптивный и полу случайного расщепления линий. Со временем классификация видов стала менее статичной и более динамичной.
Термин филогенетический , или филогения , происходит от двух древнегреческих слов ( phûlon ), означающих «раса, происхождение», и ( génesis ), что означает «происхождение, источник».
Как филогении восстанавливаются?
Филогении — это гипотезы, сформулированные на основе косвенных данных. Разбавление филогении напоминает работу следователя по раскрытию преступления, следуя следам места преступления.
Биологи часто постулируют свои филогении, используя знания из нескольких отраслей, таких как палеонтология, сравнительная анатомия, сравнительная эмбриология и молекулярная биология..
Окаменелость, хотя и неполная, дает очень ценную информацию о времени расхождения видов.
С течением времени молекулярная биология превзошла все упомянутые области, и большинство филогений выводится из молекулярных данных..
Цель реконструкции филогенетического дерева включает в себя ряд основных недостатков. Существует около 1,8 миллиона названных видов и многие другие, не описанные.
И хотя значительное число ученых ежедневно стремятся восстановить отношения между видами, у нас все еще нет полного дерева.
Гомологичные персонажи
Когда биологи хотят описать сходство между двумя структурами или процессами, они могут сделать это с точки зрения общего происхождения (гомологии), аналогий (функции) или гомоплазии (морфологического сходства)..

![Эволюция растений: огромная проблема для приверженцев эволюции [часть i]](https://lesniepolyani.ru/wp-content/uploads/1/e/7/1e71cd1a4aa4984b4acd019a3692d3ec.jpeg)
![Эволюция растений: огромная проблема для приверженцев эволюции [часть i]](https://lesniepolyani.ru/wp-content/uploads/b/c/e/bced122402abf21975f9b9621015eb63.jpeg)
![Эволюция растений: огромная проблема для приверженцев эволюции [часть i] - origins.org.ua](https://lesniepolyani.ru/wp-content/uploads/0/b/e/0be51dce8a92a8cfa302127f35123f98.webp)


Для восстановления филогении используются только гомологичные символы. Гомология является ключевым понятием в эволюции и восстановлении отношений между видами, поскольку только она адекватно отражает общее происхождение организмов..
Предположим, мы хотим сделать вывод о филогении трех групп: птиц, летучих мышей и людей. Чтобы достичь нашей цели, мы решили использовать верхние конечности как характеристику, которая помогает нам различать структуру отношений.
Поскольку птицы и летучие мыши имеют конструкции, модифицированные для полета, мы можем ошибочно сделать вывод, что летучие мыши и птицы больше связаны, чем летучие мыши, с людьми. Почему мы пришли к неправильному выводу? Потому что мы использовали аналогичный и негомологичный характер.
Чтобы найти правильные отношения, я должен искать гомологичный характер, такой как наличие волос, молочных желез и трех маленьких костей в среднем ухе — это лишь некоторые из них. Тем не менее, гомологии не легко диагностировать.
Типы
Существует три основных типа деревьев: кладограммы, аддитивные деревья и ультраметрические деревья.
Кладограммы — это простейшие деревья, которые показывают родство организмов по общему происхождению. Информация об этом типе дерева содержится в шаблонах ветвления, поскольку размер ветвей не имеет никакого дополнительного значения.
Второй тип дерева — это аддитивное дерево, также называемое метрическими деревьями или филограммами. Длина ветвей связана с количеством эволюционных изменений.
Наконец, у нас есть ультраметрические деревья или дендограммы, где все кончики деревьев находятся на одинаковом расстоянии (чего нет на филограмме, где кончик может казаться ниже или выше своего партнера). Длина ветви связана со временем эволюции.
Выбор дерева напрямую связан с эволюционным вопросом, на который мы хотим ответить. Например, если нас интересуют только отношения между людьми, кладограмма будет достаточной для исследования.
Доказательства эволюции моховидных
Среди ископаемых остатков нет никаких доказательств и эволюции моховидных (мхов, роголистников или печеночников). Следовательно, любые теории о филогенезе моховидных основываются на сравнении морфологии живых растений. Автор по фамилии Хатчинс отмечает, что моховидные существуют с древних времен, и мало изменились за это время. По словам Хатчинса, моховидные «застряли в эволюционной колее, и так там и остались». В обзоре существующей литературы по данной теме автор по фамилии Бек отмечает, что моховидные появились в самых древних слоях палеонтологической летописи и совершенно не изменились с момента своего появления.
Несогласие существует среди ученых и даже в вопросе о том, какая эволюционная дорога привела к появлению моховидном. Одни считают, что они стали связующим звеном между растениями, обитающими в воде. Другие выдвигают возражения по этому поводу, утверждая, что «палеонтологическая летопись не проливает свет на взаимоотношения и эволюционные последовательности, а факты указывают на то, что они возникли в результате «дегенерации» от такого сосудистого растения, как ринофиты». Третьи предполагают, что они эволюционировали от водорослей и наземных папоротников, а четвертые заявляют, что, скорее, моховидные эволюционировали непосредственно от водорослей. Пятые же считают, что они произошли либо от монобионтического гаплоида, либо от дибионтических зеленых водорослей. Причина, по которой возникают такие противоречия, заключается в том, что все эти теории основываются на предположениях, а не на эмпирических данных палеонтологической летописи.
У всех моховидных отсутствует водопроводящая система, а по этой причине некоторые ученые предполагают, что они стали «мостом», соединившим воду и наземные растения в процессе эволюции. Проблема, связанная с таким объяснением, заключается в том, что моховидные (в том числе мхи и печеночники) небольшие по размерам и произрастают во влажных местах. Следовательно, им не нужна такая сложная система поглощения жидкости, как ксилема и флоэма у более высоких растений. Доказательств эволюции сосудистой системы обнаружено не было, а были выявлены лишь системы, появившиеся для удовлетворения индивидуальных потребностей растений.
Поскольку современных моховидных относят к «примитивным растениям», и их также находят в очень древних слоях палеонтологической летописи, самой большой проблемой является вопрос о том, от кого они эволюционировали
По этому вопросу авторы Надакавукарен и МакКракен пришли к заключению о том, что моховидные «скорее всего, представляют собой некий эволюционный тупик, хотя адаптации, развившиеся в данной группе, были довольно успешны, принимая во внимание тот факт, что они дожили до сегодняшнего дня. Однако, по какой-то причине, мутации, которые должны были бы привести к развитию проводящей ткани, корневой системы и серозной оболочки, не проявились в генетической информации моховидных, таким образом, ограничив их в размерах и сфере распространения
Современные и ископаемые образцы очень сходны между собой, а это указывает на отсутствие изменений в пределах группы. Фактически, среда обитания всей этой группы не изменилась со времен древних моховидных».
Автор по фамилии Делевориаз делает вывод о том, что предками моховидных, возможно, были зеленые водоросли, однако не приводит на этот счет никаких доказательств, кроме указания биохимических свойств образцов живых растений, которые он исследовал.
Хотя у мхов нет развитой ксилемы или флоэмы, они обладают устьичными порами, которые регулируются замыкающими клетками эпидермиса
Тот факт, что они обладают этой системой «вентиляции», которая очень схожа с замыкающими клетками «более развитых» растений, — один из многих примеров, подтверждающих вывод РеМайна о том, что Разумный Творец посылает нам «биологическое послание», чтобы обратить внимание на Свое творение
Как филогенетические деревья интерпретируются?
Есть много способов представить филогенетическое дерево
Следовательно, важно знать, связаны ли эти различия, которые наблюдаются между двумя деревьями, с разными топологиями, то есть с реальными различиями, соответствующими двум графам, или просто с различиями, связанными со стилем представления.
Например, порядок, в котором метки появляются вверху, может варьироваться без изменения значения графического представления, обычно названия вида, рода, семейства, среди других категорий.
Это происходит потому, что деревья напоминают мобильник, где ветви могут вращаться, не изменяя отношения представленных видов.
В этом смысле не имеет значения, сколько раз изменяется порядок или вращаются «зависшие» объекты, поскольку это не меняет способ их соединения — и это важно
Аннотация
Из чего состоят живые тела и при чем тут углерод? Что такое генетический код, кто такие вирусы, как устроено эволюционное древо и почему произошел кембрийский взрыв? Предлагаемая книга дает актуальные ответы на эти и многие другие вопросы. «Фокусом» рассказа служит эволюция жизни на Земле: автор считает, что только под этим углом зрения самые разные биологические проблемы обретают единый смысл. Книга состоит из четырех частей, темы которых последовательно расширяются: «Химия жизни», «Механизм жизни», «Древо жизни» и «История жизни».
![Эволюция растений: огромная проблема для приверженцев эволюции [часть i] - origins.org.ua](https://lesniepolyani.ru/wp-content/uploads/c/b/4/cb4869e13763e821147995b856f330bc.jpeg)
![Эволюция растений: огромная проблема для приверженцев эволюции [часть i]](https://lesniepolyani.ru/wp-content/uploads/8/7/e/87ec666652e569e7233595b32c0ae18c.jpeg)



![Классификация и филогения цветковых растений (а. л. тахтаджян) [1980 - - жизнь растений. том 5. часть 1. цветковые растения]](https://lesniepolyani.ru/wp-content/uploads/2/7/7/277afd92bd0e272a8296eb2124ccad47.jpeg)
![Эволюция растений: огромная проблема для приверженцев эволюции [часть i] - origins.org.ua](https://lesniepolyani.ru/wp-content/uploads/b/0/0/b007daefdd7e45af085bee5464f3382a.jpeg)
![Эволюция растений: огромная проблема для приверженцев эволюции [часть i] - origins.org.ua](https://lesniepolyani.ru/wp-content/uploads/f/3/f/f3f8a12e9ecb17901411c291c552d3d0.webp)
![Классификация и филогения цветковых растений (а. л. тахтаджян) [1980 - - жизнь растений. том 5. часть 1. цветковые растения]](https://lesniepolyani.ru/wp-content/uploads/5/c/a/5ca4a2c15f00c1d121a2708a7b2851e0.jpeg)
Рекомендуется широкому кругу читателей, всерьез интересующихся современной биологией.
Книга выходит в серии Primus при поддержке фонда «Эволюция» и программы «Книжные проекты Дмитрия Зимина».
Скачать или читать онлайн книгу От атомов к древу. Введение в современную науку о жизни
На этой странице свободной электронной библиотеки fb2.top вы можете ознакомиться
с описанием книги «От атомов к древу. Введение в современную науку о жизни» и другой информацией о ней, а затем начать
читать книгу онлайн с помощью читалок, предлагаемых по ссылкам под постером, или
скачать книгу в формате fb2 на свой смартфон, если вам больше по вкусу сторонние
читалки. Книга написана автором Сергей Александрович Ястребов,
является частью серий: Библиотека фонда «Эволюция»,
Книжные проекты Дмитрия Зимина,
Primus,
относится к жанрам Химия,
Биология, биофизика, биохимия,
Научная литература,
Зарубежная образовательная литература, зарубежная прикладная, научно-популярная литература,
добавлена в библиотеку 21.10.2018.
С произведением «От атомов к древу. Введение в современную науку о жизни» , занимающим объем 542 печатных страниц,
вы наверняка проведете не один увлекательный вечер. В онлайн читалках,
которые мы предлагаем посетителям библиотеки fb2.top, предусмотрен ночной режим
чтения, который отлично подойдет для тёмного времени суток и чтения перед сном.
Помимо этого, конечно же, можно читать «От атомов к древу. Введение в современную науку о жизни» полностью в
классическом дневном режиме или же скачать книгу на свой смартфон в удобном
формате fb2. Желаем увлекательного чтения!
С этой книгой читают:
-
Люди мира. Русское научное зарубежье
— Сергей Ястребов -
Предчувствие «шестой волны»
— Сергей Ястребов -
Кислород. Молекула, изменившая мир
— Ник Лэйн -
На качелях XX века
— Александр Несмеянов -
Немые свидетели
— Владимир Санов, Михаил Любарский -
Химия, изменившая мир
— Роман Потапов -
Загадка «Таблицы Менделеева»
— Петр Дружинин -
Эта увлекательная химия
— Георгий Шульпин -
Чудесная молекула
— Лионель Салем -
Квантовая химия — ее прошлое и настоящее. Развитие электронных представлений о природе химической связи
— Сергей Семенов, Игорь Дмитриев
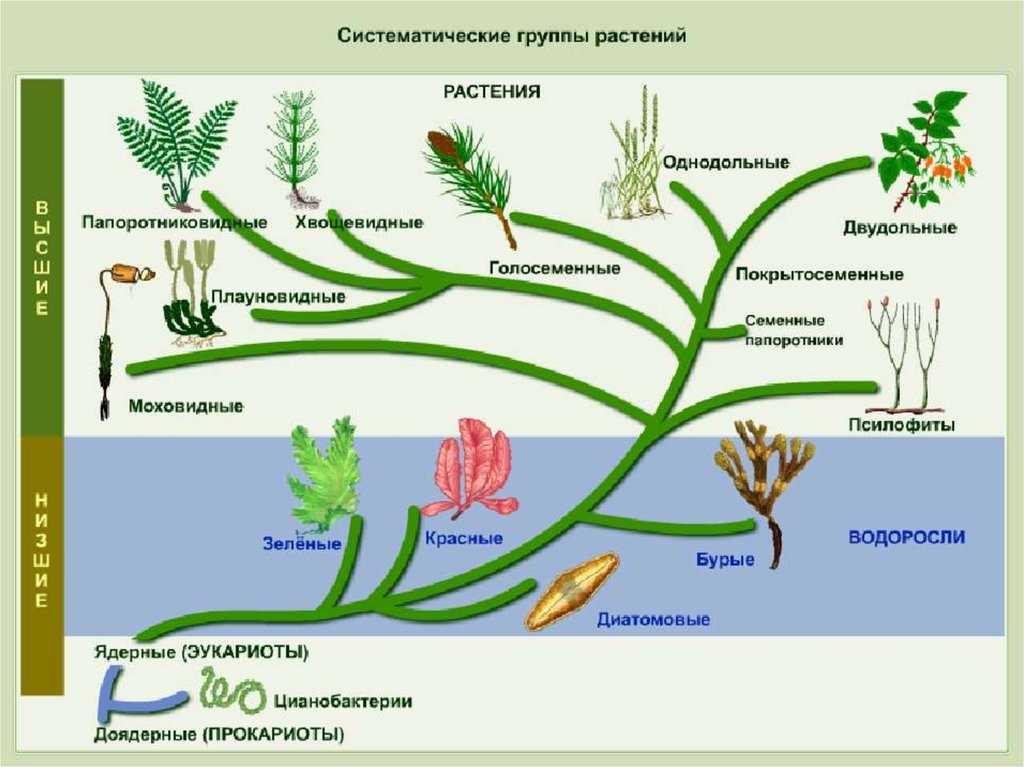
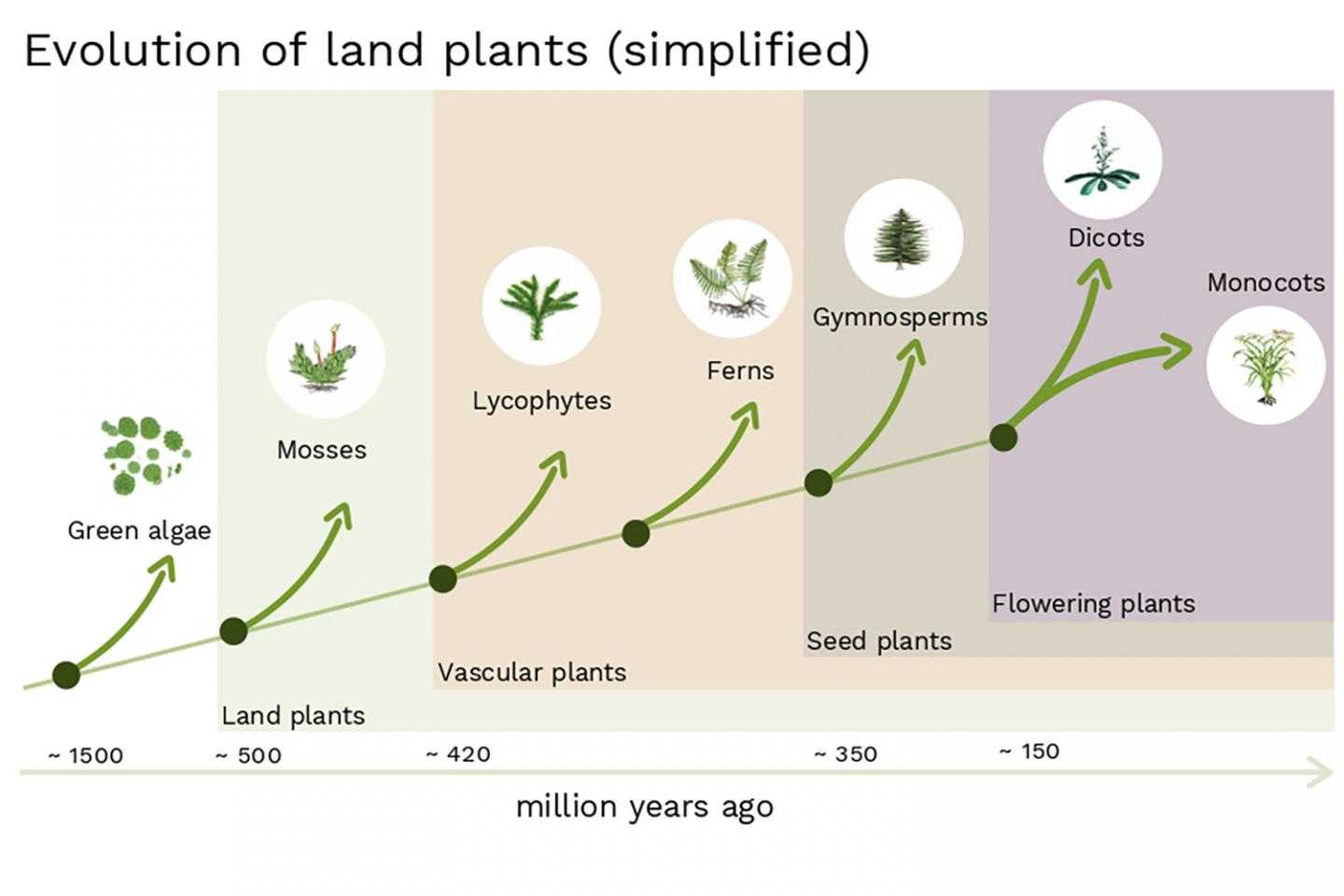
Филогенетическое дерево растений
Еще более сложной задачей для эволюционистов стала разработка филогенетического дерева для растений (по сравнению с филогенетическим деревом животного мира). И это объясняется несколькими причинами. Только за последнее десятилетие было опубликовано как минимум 15 вариантов филогенетических деревьев цветковых растений. Одна из главных проблем – сложность разработки естественной схемы классификации, которая распределяет «растения по степени примитивности или продвинутости. Для этого нужно было бы определить характеристики, которые следует считать примитивными или продвинутыми. Как мы увидим далее, определить это не так уж просто. А в большинстве случаев, из-за полного отсутствия доказательств, определить это просто невозможно
Перед каждым из нас стоит выбор – либо довольствоваться неполной системой, либо произвольно принимать решения о важности определенных характеристик. Факт в том, что такая неполная система не удовлетворит никого, поэтому неизбежно приходится принимать произвольные решения. Но как только такое произвольное решение войдет в схему классификации, сама эта система станет в такой же степени искусственной»
Но как только такое произвольное решение войдет в схему классификации, сама эта система станет в такой же степени искусственной».
Как следует из приведенной ниже цитаты, эволюционисты вынуждены ссылаться на существующую палеонтологическую летопись, а также на известные сегодня растения, чтобы делать предположения о том, что из чего эволюционировало.
«На сегодняшний день принято считать, что покрытосеменные эволюционировали из неких примитивных голосеменных, возможно из кустарников. Возможных вариантов в меловом периоде не выявлено, однако в более ранних мезозойском и палеозойском периодах присутствуют голосеменные, в которых проявляются определенные характеристики, сходные с признаками покрытосеменных. Это само по себе свидетельствует о более раннем происхождении покрытосеменных, по сравнению с появлением палеонтологической летописи».
Эволюционная классификация
Монофилетические линии
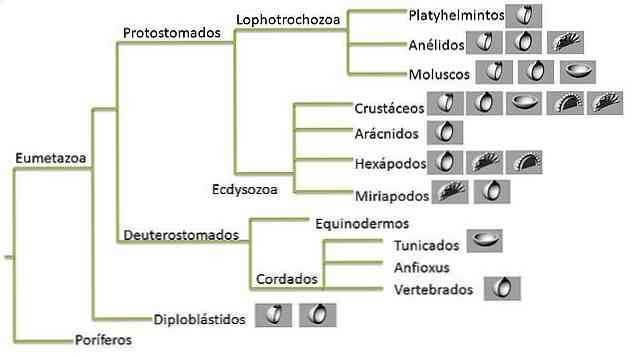
Эволюционные биологи стремятся найти классификацию, которая соответствует паттерну ветвления филогенетической истории групп. В этом процессе был разработан ряд терминов, широко используемых в эволюционной биологии: монофилетический, парафилетический и полифилетический..
Таксон или монофилетическая линия — это та, которая включает в себя нецентральный вид, который представлен в узле, и всех его потомков, но не другие виды. Эта группировка называется кладой.
Монофилетические линии определяются на каждом уровне таксономической иерархии. Например, семейство Felidae, линия, которая содержит кошек (включая домашних кошек), считается монофилетической..
Аналогичным образом Animalia также является монофилетическим таксоном. Как мы видим, семья Felidae находится в Animalia, поэтому монофилетические группы могут быть вложенными.
Парафилетические и полифилетические линии
Однако не все биологи разделяют мысль о кладистической классификации. В тех случаях, когда данные не являются полными или просто для удобства, называются определенные таксоны, которые включают виды разных клад или более высоких таксонов, которые не имеют более общего общего предка.
Таким образом, полифилетический таксон определяется как группа, включающая организмы разных клад, которые не имеют общего предка. Например, если мы хотим обозначить группу гомеотерм, это будет включать птиц и млекопитающих.
Напротив, парафилетическая группа не содержит всех потомков самого последнего общего предка. Другими словами, исключите любого из членов группы. Наиболее часто используемым примером являются рептилии, эта группа не содержит всех потомков самого последнего общего предка: птиц.
Что такое филогенетическое дерево?
Филогенетическое древо — это графическое представление эволюционной истории группы организмов. Эта модель исторических отношений — филогения, которую исследователи пытаются оценить.
Деревья состоят из узлов, которые соединяются с «ветвями». Конечные узлы каждой ветви представляют собой терминальные таксоны и представляют последовательности или организмы, для которых известны данные — это могут быть живые или вымершие виды.
Внутренние узлы представляют гипотетических предков, в то время как предок, найденный в корне дерева, представляет предка всех последовательностей, представленных в графе..
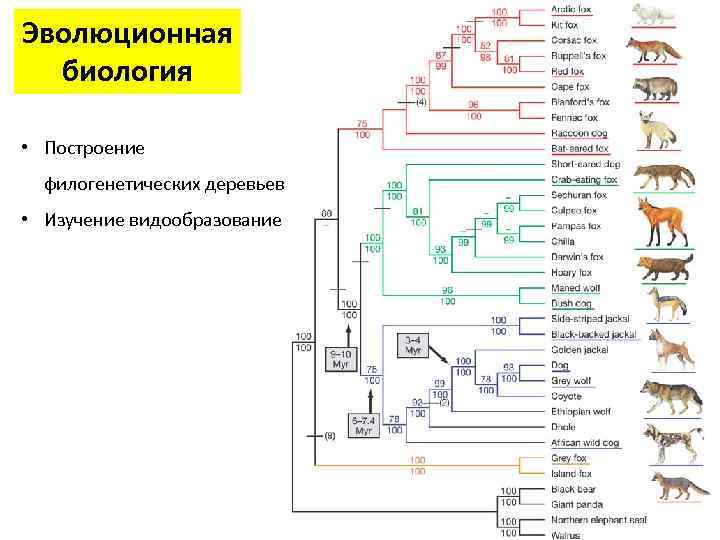
Строительство
Филогенетические деревья, состоящие из нетривиального числа входных последовательностей, строятся с использованием методов вычислительной филогенетики . Методы Расстояние-матрицы , такие как сосед-присоединения или UPGMA , которые вычисляют генетическое расстояние от множественного выравнивания последовательностей , наиболее просты в реализации, но не вызывают эволюционной модели. Многие методы выравнивания последовательностей, такие как ClustalW, также создают деревья с помощью более простых алгоритмов (то есть основанных на расстоянии) построения дерева. Максимальная экономия — еще один простой метод оценки филогенетических деревьев, но подразумевает неявную модель эволюции (то есть экономию). Более продвинутые методы используют критерий оптимальности по максимальной вероятности , часто в рамках байесовского и применить явную модель эволюции оценки филогенетического дерева. Идентификация оптимального дерева с использованием многих из этих методов является NP-сложной задачей , поэтому методы эвристического поиска и оптимизации используются в сочетании с функциями оценки дерева для определения достаточно хорошего дерева, которое соответствует данным.
Методы построения деревьев можно оценить по нескольким критериям:
- эффективность (сколько времени нужно, чтобы вычислить ответ, сколько памяти ему нужно?)
- мощность (хорошо ли используются данные или информация тратится зря?)
- согласованность (будет ли он постоянно сходиться к одному и тому же ответу, если каждый раз давать разные данные для одной и той же модельной задачи?)
- устойчивость (хорошо ли справляется с нарушениями предположений базовой модели?)
- фальсифицируемость (предупреждает ли она нас о том, что использование нецелесообразно, т.е. когда предположения нарушаются?)
Методы построения деревьев также привлекли внимание математиков. Деревья также могут быть построены с использованием Т-теории .
Форматы файлов
Деревья могут быть закодированы в различных форматах, каждый из которых должен представлять вложенную структуру дерева. Они могут кодировать или не кодировать длины ветвей и другие особенности. Стандартизированные форматы критически важны для распространения и совместного использования деревьев, не полагаясь на вывод графики, который трудно импортировать в существующее программное обеспечение. Обычно используемые форматы:

![Классификация и филогения цветковых растений (а. л. тахтаджян) [1980 - - жизнь растений. том 5. часть 1. цветковые растения]](https://lesniepolyani.ru/wp-content/uploads/b/8/4/b84179c2552f64f57c897519de91c2d6.jpeg)
![Эволюция растений: огромная проблема для приверженцев эволюции [часть i]](https://lesniepolyani.ru/wp-content/uploads/5/0/3/50373bd14dcaf4037c9d910154c652fa.jpeg)
![Эволюция растений: огромная проблема для приверженцев эволюции [часть i] - origins.org.ua](https://lesniepolyani.ru/wp-content/uploads/3/7/3/37340e16b2bb3de5c33582bc35318a8a.jpeg)
- Формат файла Nexus
- Формат Ньюика
Что такое филогения?
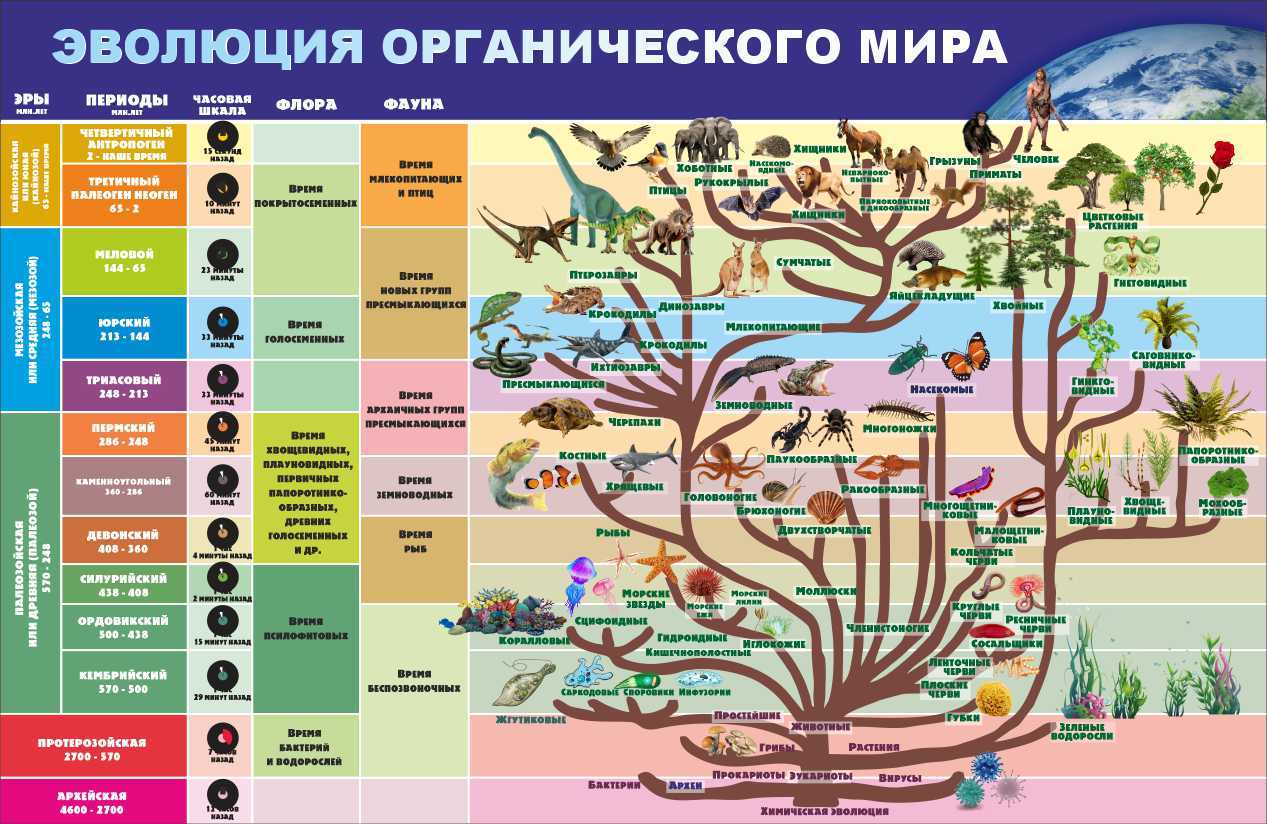
В свете биологических наук, одним из самых удивительных событий, которые произошли, является эволюция. Указанное изменение органических форм с течением времени может быть представлено в филогенетическом дереве. Таким образом, филогения выражает историю происхождения и как они изменились с течением времени.
Одним из прямых следствий этого графа является общее происхождение. То есть, все организмы, которые мы видим сегодня, появились как потомки с модификациями прошлых форм. Эта идея была одной из самых значительных в истории науки.
Все формы жизни, которые мы можем оценить сегодня — от микроскопических бактерий до растений и более крупных позвоночных — связаны, и эти отношения представлены в огромном и сложном древе жизни.
По аналогии с деревом, виды, которые сегодня живут, будут представлять листья, а остальные ветви — их эволюционную историю..
Дополнительная терминология
Помимо этих трех основных понятий, существуют и другие необходимые термины, когда речь идет о филогенетических деревьях:
Политомия: когда у филогенетического дерева более двух ветвей в узле, говорят, что существует политомия. В этих случаях филогенетическое древо не разрешено полностью, потому что отношения между вовлеченными организмами не ясны. Обычно это происходит из-за отсутствия данных и может быть исправлено только тогда, когда исследователь накапливает больше.
Внешняя группа: в филогенетических темах часто можно услышать понятие внешней группы, также называемое вне группы. Эта группа выбрана, чтобы иметь возможность корневать дерево. Его следует выбрать как таксон, ранее отделившийся от исследуемой группы. Например, если я изучаю иглокожих, вы можете использовать аутгруппа морские брызги.
Мы не можем сделать вывод о существовании нынешних предков или «старых» видов.
Когда мы будем говорить о видах текущий мы не должны применять к ним наследственные коннотации. Например, когда мы думаем об отношениях между шимпанзе и людьми, мы можем неправильно понять, что шимпанзе являются предками нашей линии.
Однако общего предка шимпанзе и человека не было. Думать, что шимпанзе — это предки, значило бы предполагать, что его эволюция прекратилась, когда обе линии разделились.
Следуя той же логике этих идей, филогенетическое дерево также не сообщает нам, есть ли молодые виды. Поскольку частоты аллелей постоянно меняются, и со временем меняются новые признаки, трудно определить возраст вида, и, конечно же, дерево не дает нам такой информации.
«Сдвиг частот аллелей во времени» — это то, как популяционная генетика определяет эволюцию.