

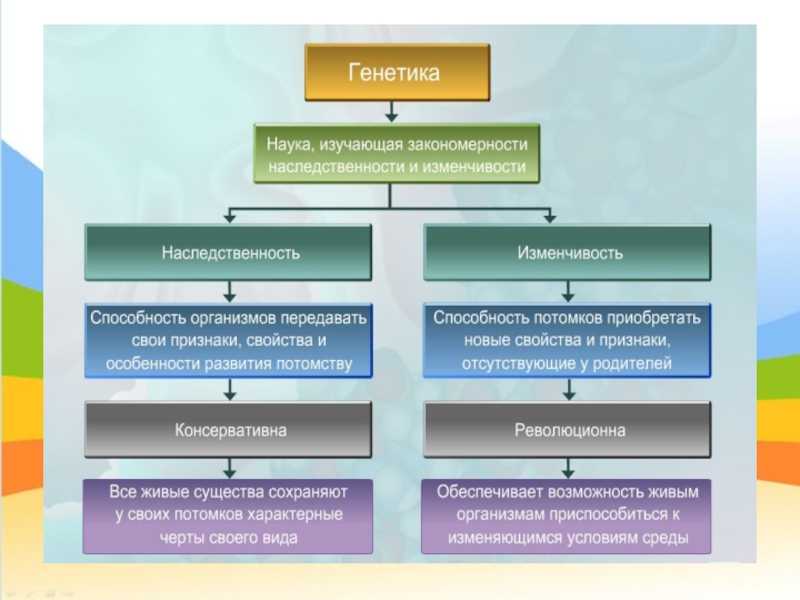
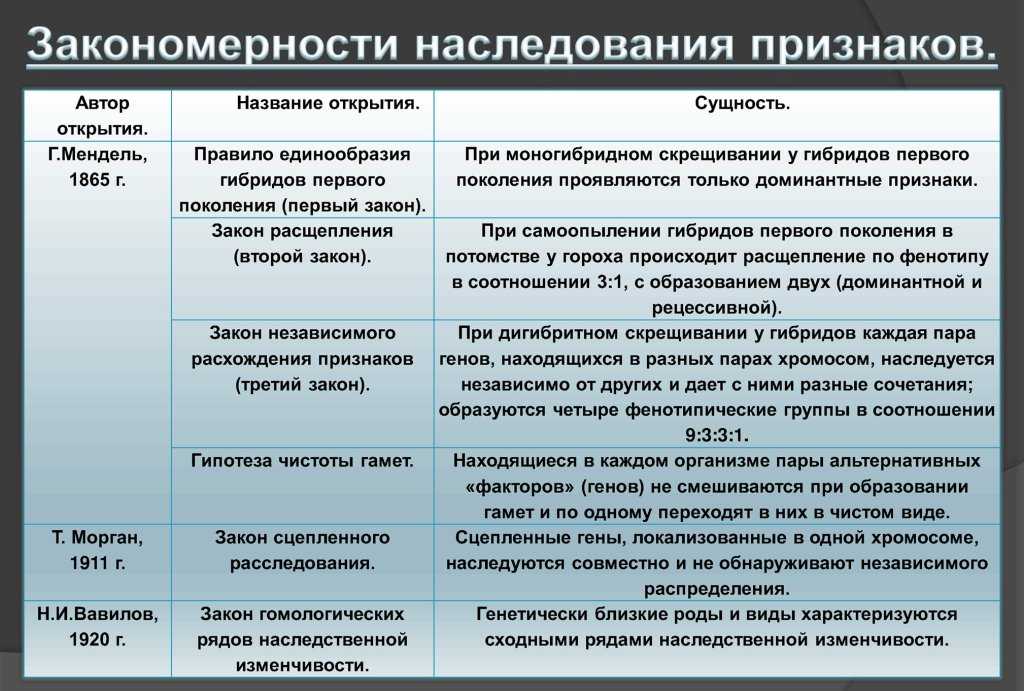
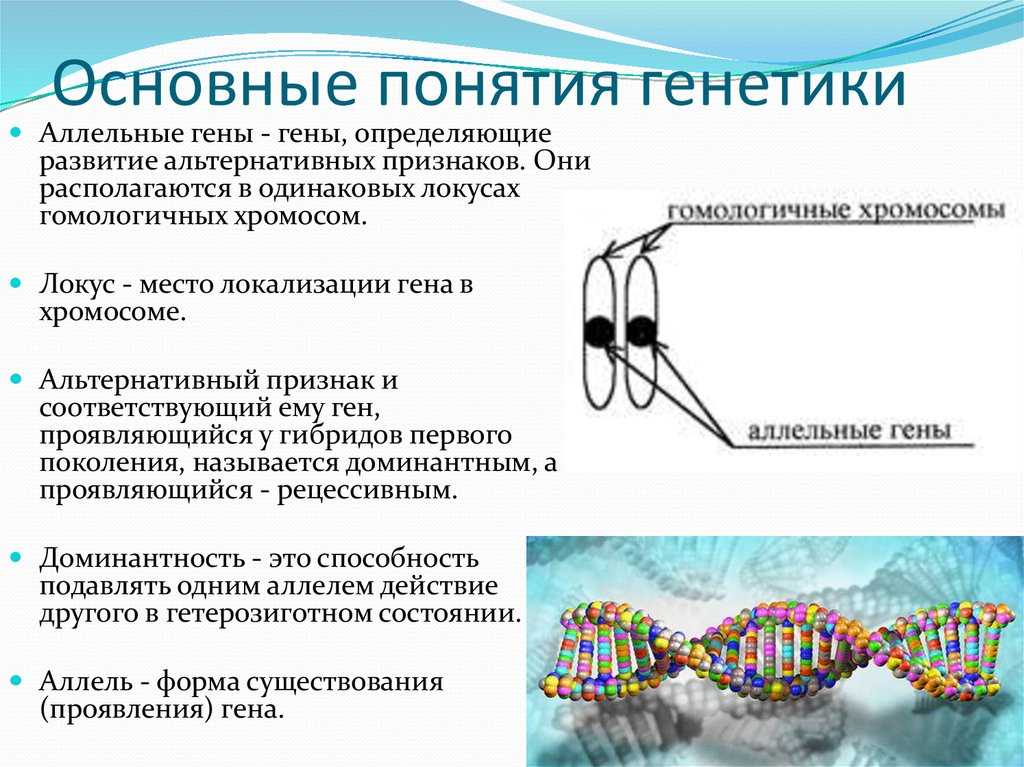
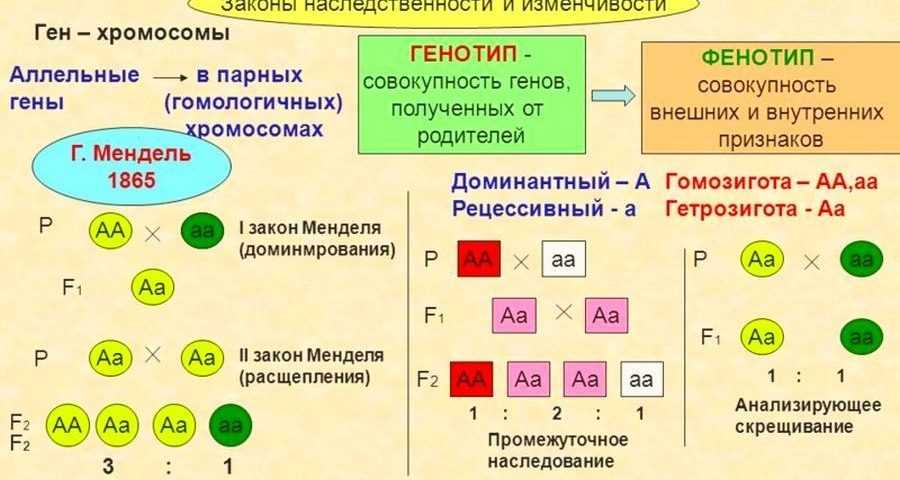
Сформулированные Г. Менделем законы
Правило доминирования, или первый закон: при моногибридном скрещивании у гибридов первого поколения проявляются только доминантные признаки — оно фенотипически единообразно.
Закон расщепления, или второй закон Г. Менделя: при скрещивании гибридов первого поколения в потомстве происходит расщепление признаков в отношении 3:1 — образуются две фенотипические группы — доминантная и рецессивная.
Закон независимого наследования (третий закон): при дигибридном скрещивании у гибридов каждая пара признаков наследуется независимо от других и дает с ним разные сочетания. Образуются четыре фенотипические группы, характеризующиеся отношением 9:3:3:1.
Ход моногибридного скрещивания (первый и второй законы Менделя)
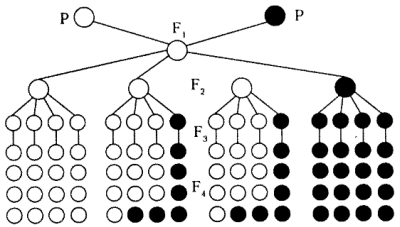
Светлые кружки — организмы с доминантными признаками; темные — с рецессивным признаком.
| Правило доминирования, или первый закон | В первом поколении F1, в результате скрещивания гороха с желтыми и зелеными семенами все семена оказываются желтыми (единообразными) |
| Закон расщепления, или второй закон Менделя | в F2 наблюдается расщепление в соотношении 3:1, то есть три четверти особей несут доминантный признак и одна четверть — рецессивный |
Гипотеза чистоты гамет: находящиеся в каждом организме пары альтернативных признаков не смешиваются и при образовании гамет по одному от каждой пары переходят в них в чистом виде.
Для объяснения наблюдаемых закономерностей Мендель выдвинул гипотезу чистоты гамет, предположив следующее:
- любой признак формируется под влиянием материального фактора (гена).
- Фактор, определяющий доминантный признак, он определил заглавной буквой А, а рецессивный — а. Каждая особь содержит два фактора, определяющих развитие признака, один она получает от матери, другой — от отца.
- При образовании гамет у животных и спор — у растений происходит редукция факторов и в каждую гамету или спору попадает только один.
Согласно этой гипотезе ход моногибридного скрещивания записывают так:
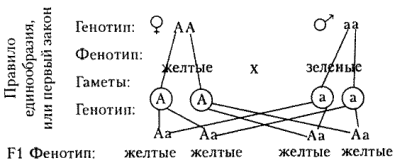
При любых сочетаниях гамет все гибриды имеют одинаковый генотип и фенотип.
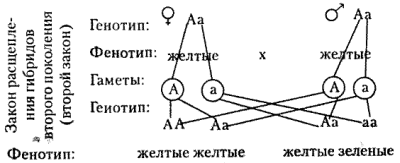
В F2 расщепление по генотипу будет 1АА; 2Аа; 1аа, а но фенотипу: 3 желтых, 1 зеленый (3:1).
Иногда у гибридов F1, не наблюдается полного доминирования, их признаки носят промежуточный характер. Такой характер наследования называют промежуточным, или неполным доминированием.
Пример: моногибридное скрещивание ночной красавицы: при неполном доминировании в F2 расщепление по фенотипу и генотипу выражается одинаковым соотношением: 1:2:1 (1 белый, 2 розовых, 1 красный).
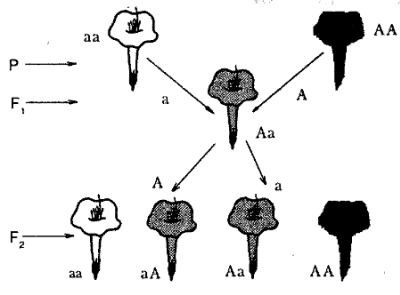
Дигибридное скрещивание
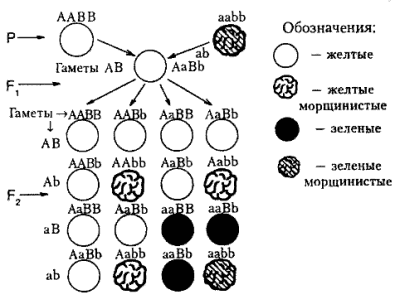
Здесь гомозиготные родители отличались друг от друга по двум парам признаков; окраска семян (желтая и зеленая), форме семян (гладкая и морщинистая). В F2 произошло расщепление в соотношении 9:3:3:1.
Характер наследования был определен как независимое и сформулирован третий закон Менделя, или закон независимого наследования.
Независимое наследование имеет огромное значение для эволюции, так как является источником комбинативной изменчивости и многообразия живых организмов.
Расщепление признаков при скрещивании
Полное и неполное доминирование как генное взаимодействие происходит в соответствии с арифметикой законов Г. Менделя. В первом случае соотношение в F2 фенотипов (3:1) не совпадает с соотношением генотипов потомков (1:2:1), поскольку фенотипически сочетания аллелей АА
и Аа
проявляются одинаково. Тогда неполное доминирование — это совпадение в F2 пропорции отличающихся генотипов и фенотипов (1:2:1).
У земляники окраска я год наследуется по принципу неполного доминирования. Если скрестить растение с красными ягодами (АА
) и растение с белыми ягодами — генотип аа
, то в первом поколении все полученные растения дадут плоды с розовой окраской (Аа
).
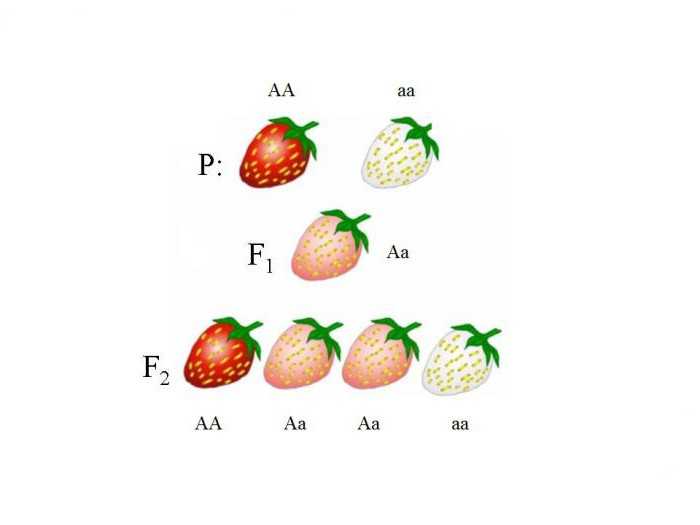
Скрестив гибриды из F1, во втором поколении F2 получим соотношение потомков, совпадающее с таковым генотипов: 1АА+2Аа+1аа
. По 25% растений из второго поколения дадут красные и неокрашенные плоды, 50% растений — розовые.
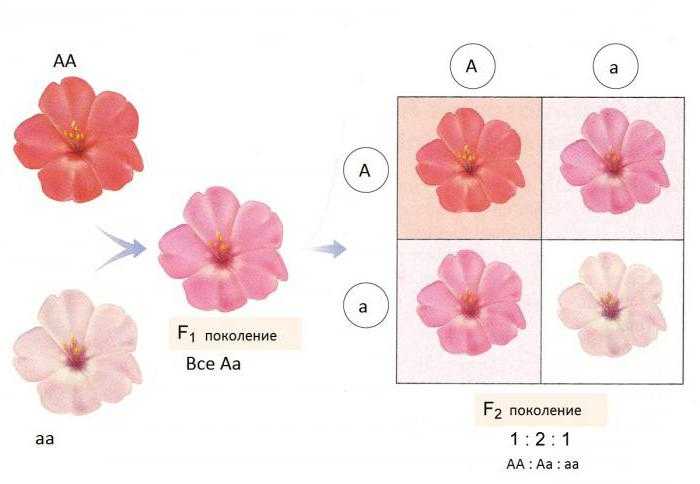
Аналогичную картину будем наблюдать в двух поколениях при скрещивании чистых линий цветков ночной красавицы с венчиками пурпурного и белого цветов.
Доминирование генов
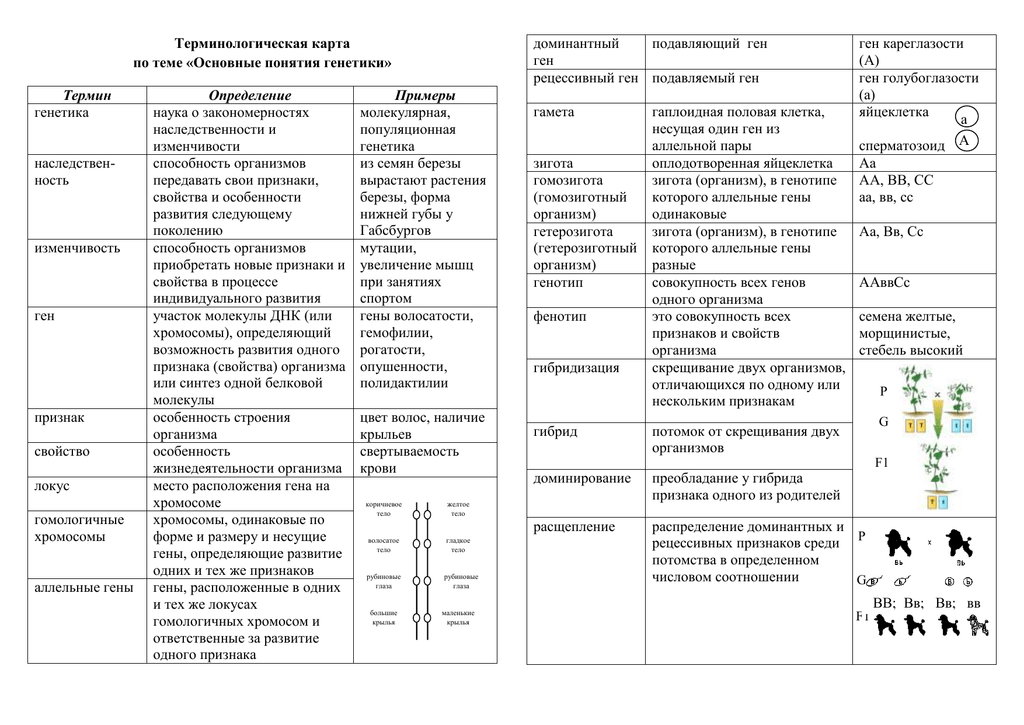
Вы когда-нибудь задумывались, почему у вас есть такой особый цвет глаз или тип волос? Все это связано с передачей генов. Как обнаружил Грегор Мендель, черты наследуются при помощи передачи генов от родителей к их потомкам. Гены представляют собой участки ДНК, расположенные на наших хромосомах. Они передаются от одного поколения к другому через половое размножение. Ген для конкретного признака может существовать в более чем одной форме или аллеле. Для каждой характеристики или признака клетки животных обычно наследуют два аллеля. Парные аллели могут быть гомозиготными (с идентичными аллелями) или гетерозиготными (с разными аллелями) для данного признака.
Когда пары аллелей одинаковы, генотип этого признака идентичен, а фенотип или характеристика, которая наблюдается, определяется гомозиготными аллелями. Когда парные аллели для признака различные или гетерозиготные, возможно несколько вариантов. Гетерозиготные аллели, которые обычно наблюдаются в клетках животных, включают полное доминирование, неполное доминирование и кодоминирование.
Закон сцепленного наследования
В 1911 году Томасом Морганом был сформулирован закон сцепленного наследования — сцепленные гены, локализованные в одной хромосоме, наследуются вместе и не обнаруживают независимого расщепления.
В каждой хромосоме сосредоточено несколько тысяч генов, по которым одна особь данного вида отличается от другой. Выясняя вопрос, как будут наследоваться признаки этих генов, Морган установил, что гены, расположенные в одной хромосоме, наследуются сцеплено, вместе, как одна альтернативная пара, не обнаруживая независимого наследования.
Сцепление не всегда бывает абсолютным. В профазе первого деления мейоза при конъюгации хромосом происходит их перекрест, вследствие чего гены, находящиеся в одной хромосоме, оказывались в разных гомологических хромосомах и попадали в разные гаметы.

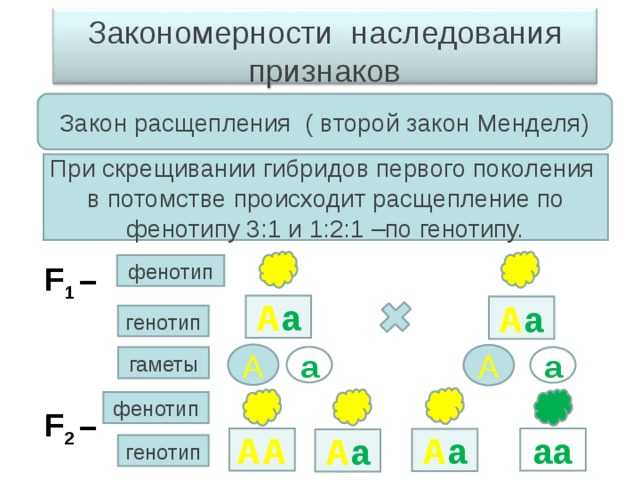



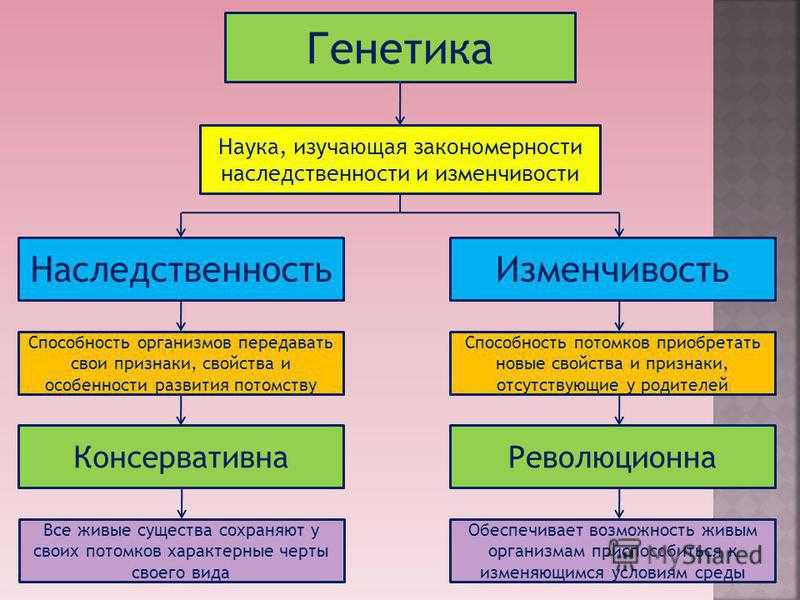



Схема перекреста хромосом
Два гена, расположенные в одной хромосоме (светлые круги в одной из хромосом), в результате перекреста оказываются в разных гомологичных хромосомах.
Такой обмен приводит к перегруппировке сцепленных генов и является одним из источников комбинативной изменчивости.
Перекрест хромосом играет определенную роль в эволюции, так как новое сочетание генов вызывает появление новых признаков, которые могут оказаться полезными или вредными для организма и повлиять на их выживаемость.
Ген может одновременно влиять на формирование нескольких признаков, проявляя при этом множественное действие.
Наследование коричневой и платиновой окраски меха у норок
Задача 375.
На ферме разводят норок коричневой и платиновой окраски меха. Самку с коричневым мехом скрестили с платиновым самцом. Какими будут их потомки первого и второго поколений по генотипу и фенотипу, если ген платиновой окраски меха – рецессивный?Решение:
«+» — коричневый мех (дикий тип окраса);
р — платиновый мех.
Схема скрещивания (1)
Р: «+»»+» х рр
Г: «+» р
F1: «+»p — 100%.Фенотип:
«+»p — коричневая норка — 100%.
Наблюдается единообразие первого поколения.
Схема скрещивания (2)
Р: «+»p х «+»p
Г: «+»; p «+»; p
F2: «+»»+» — 25%; «+»p — 50%; pp — 25%.
Наблюдается 3 типа генотипа. Расщепление по генотипу — 1:2:1.Фенотип:
«+»»+» — коричневая норка — 25%;
«+»p — коричневая норка — 50%;
pp — платиновая норка — 25%.Наблюдаемый фенотип:
коричневая норка — 75%;
платиновая норка — 25%.
Наблюдается 2 типа фенотипа. Расщепление по фенотипу — 3:1.
Молекулярные механизмы [ | ]
Молекулярные основы доминирования были неизвестны Менделю . В настоящее время ясно, что локус , соответствующий определённому гену , состоит из длинных последовательностей, включающих сотни и тысячи нуклеотидов ДНК . Центральная догма молекулярной биологии гласит, что ДНК → РНК → белок , то есть ДНК транскрибируется в мРНК , а мРНК транслируется в белок. В этом процессе различные аллели могут транскрибироваться или не транскрибироваться, а будучи затранскрибированными, транслироваться в различные формы одного и того же белка — изоформы . Часто белки функционируют как ферменты , катализирующие химические реакции в клетке , которые прямо или косвенно определяют фенотип . У любого диплоидного организма аллели, соответствующие одному локусу, являются либо одинаковыми (у гомозигот), либо разными (у гетерозигот). Даже если на уровне последовательностей ДНК аллели различны, то их белки могут быть идентичными. В отсутствие различий между белковыми продуктами невозможно сказать, какой из аллелей доминирует (в этом случае имеет место кодоминирование). Даже если два белковых продукта слегка отличны друг от друга, они, вероятно, дают одинаковый фенотип и могут осуществлять одинаковые ферментативные реакции (если они являются ферментами). В этом случае также невозможно сказать, какой из аллелей доминирует.
Доминирование, как правило, возникает, когда один из аллелей является нефункциональным на молекулярном уровне, то есть не транскрибируется или даёт нефункциональный белковый продукт. Это может быть результатом мутации, изменяющей последовательность ДНК аллеля. У гомозиготы по нефункциональным аллелям, как правило, проявляется характерный фенотип из-за отсутствия определённого белка. Например, у людей и других животных непигментированная кожа альбиносов проявляются из-за гомозиготности по аллелю, препятствующему синтезу кожного пигмента меланина
Важно понимать, что рецессивность определяется у аллеля не по отсутствию какой-либо функции: у гетерозигот это является результатом взаимодействия с альтернативным аллелем. Возможны три основных типа таких взаимодействий:
Неполное доминирование в генетике. Кодоминирование в генных аллелях.
В одном из скрещиваний, которые проводил Мендель, доминантный признак не полностью исключал проявление рецессивного признака. При скрещивании крупнолистного сорта гороха с мелколистным в первом поколении наблюдался промежуточный фенотип: листья имели среднюю величину. В последующих экспериментах выяснилось, что, обусловленный неполным доминированием одного аллеля над другим промежуточный фенотип у гибридов первого поколения встречается у различных организмов.
При неполном доминировании могут наблюдаться некоторые вариации признака у гибридов с отклонениями в сторону доминантного или рецессивного фенотипа. Если выражение признака у гетерозигот примерно промежуточное, то такие признаки называются полудоминантными. Так у львиного зева (Antirrhinum majus) и ночной красавицы (Mirabilis jalapa) гибриды от скрещивания красно цветковых растений с белоцветковыми имеют розовую окраску. Другим примером полудоминантного признака может служить окраска оперения у кур андалузской породы.
При скрещивании белых кур с черными петухами, потомство F, имеетсерую окраску, вто время как во втором поколении у 1/4 птиц окраска оперения черная, у 1/2 серая и у 1/4 белая.
Кодоминирование в генных аллелях.
В случае кодоминирования у гетерозигот полностью проявляются оба аллеля. Наиболее яркий пример кодоминирования — наследование групп крови АВО у человека. Группы крови О (I), А (II), В (III), АВ (IV) детерминируются геном / (для обозначения генов принят курсивный шрифт). Известны три основных аллеля этого гена, два из которых IA и IB доминантные, а один I0 — рецессивный. При гомозиготности IАIА эритроциты имеют только поверхностный антиген А (группа крови А).
При гомозиготности IвIв эритроциты несут другой антиген — В (группа крови В). В случае гомозиготности I0I0 эритроциты лишены обоих антигенов (группа крови О). У гетерозигот IАI0 или IBI0 в соответствии с имеющимся антигеном группа крови А или В. У гетерозигот IAIB эритроциты несут оба антигена А и В (гpyппa крови АВ).
Как правило, на уровне синтеза полипептидов аллели гетерозигот кодоминантны. Так у гетерозигот по серповидноклеточной анемии HbA/HbS, гемоглобины HbS составляют 35-42%, НbА 60-65%. Полная клиническая картина заболевания (тромбозы мелких сосудов, инфаркты внутренних органов, гемолитическая анемия, гиперплазия костного мозга, нарушения мозгового кровообращения; смерть в возрасте 3-10 лет, иногда позже) проявляется только в случае гомозиготности по HbS. У гомозигот мутантный гемоглобин Hb состоит из двух нормальных цепей и двух мутантных бета-цепей, в 6-м положении которых глутаминовая кислота заменена валином.
Заболевание наследуется по аутосомно-рецессивному типу, наличие у гетерозигот до 65% нормального гемоглобина позволяет большинству из них чувствовать себя здоровыми. Однако в холодное время года, при повышенной нагрузке, а также при полетах на самолете у них появляются боли в суставах, сердце, брюшной полости, в области селезенки, что свидетельствует о неполной доминантности нормального аллеля НbА в экстремальных условиях у носителей мутантного аллеля HbS. Таким образом, кодоминантность у гетерозигот HbA/HbS проявляется только на уровне синтеза полипептидных цепей гемоглобина, а заболевание с полной клинической картиной наследуется по аутосомно-рецессивному типу.
Кодоминантность
Для нашего примера кодоминирования предположим, что мы скрещиваем коров, у которых есть правила кодоминирования для их окраса шерсти. Коровы с генотипом BB полностью черные, с генотипом WW полностью белые, и при скрещивании коровы с генотипом BW имеют черные и белые пятна по всему телу. (При скрещивании, которое следует моделям наследования кодоминирования, все прописные буквы обычно используются для обозначения аллелей, чтобы показать, что ни один аллель не доминирует над другим.)
К настоящему времени вы, вероятно, можете сказать, что если бы вы скрестили чистую черную корову с чисто белой, у всех потомков были бы черные и белые пятна, поскольку все они имели генотип BW.
Ниже показан квадрат Пеннета, показывающий, что происходит, когда вы скрещиваете чисто черную корову (BB) с черно-белой пятнистой коровой (BW).
Из квадрата Паннета видно, что половина потомства будет чисто черной, а другая половина — с черными и белыми пятнами.
Примеры неполного доминирования
Неполное доминирование может встречаться у представителей всех живых организмов. У человека доминантными признаками являются курчавость волос, карий цвет глаз, длинные ресницы, веснушки и близорукость.
К примеру, при наследовании структуры волос ген курчавых волос доминирует над геном прямых волос не в полной мере. Тогда промежуточным признаком станут волнистые волосы, которые унаследует первое поколение.
По законам неполного доминирования передаются и наследственные заболевания. Болезнь Тея-Сакса, поражающая нервную систему, носит аутосомно-рецессивный характер. Люди, страдающие этим заболеванием, имеют два рецессивных гена, которые его вызывают. Но один или оба родителя могли быть носителями рецессивных генов, вырабатывающих половину необходимого для нормальной жизни фермента.
Наиболее широко изучается неполное доминирование у домашних животных. Задачей исследований является выведение здоровых животных, несущих определенную ценность.
При скрещивании короткошерстного кролика рекса и длинношерстного ангорского кролика получается кролик со средней длиной шерсти, отличной от длины шерсти любого из родителей.
Пара лошадей с красным аллелем у одного из родителей и кремовым аллелем у другого даст потомство с золотистым цветом шерсти (паломино или оленья шкура).
Более одиннадцати тысяч лет назад человек начал заниматься сельским хозяйством. Все это время происходит скрещивание растений по определенным признакам для повышения качества и количества урожая.








Кукуруза с несколькими рецессивными генами является более здоровой и дает лучший урожай, чем растения с доминантными генами. Предок современной кукурузы теосинте содержит в початке всего 9-12 зерен.
Еще одним примером неполного доминирования у растений является цвет баклажанов. При скрещивании темно-фиолетовых и белых сортов получаются плоды светло-фиолетового цвета.
В изучении генетики явление неполного доминирования играет важную роль. Решением вопросов повышения урожайности или выведения здорового потомства становится создание нового фенотипа с набором определенных характеристик.
Относительный характер доминирования [ | ]
Как уже отмечалось выше, характер доминирования зависит от уровня анализа признака. Рассмотрим это на примере серповидно-клеточной анемии . Гетерозиготные носители гена гемоглобина S (AS) на уровне моря имеют нормальную форму эритроцитов и нормальную концентрацию гемоглобина в крови (полное доминирование А над S). На больших высотах (более 2,5-3 тыс. м) у гетерозигот концентрация гемоглобина понижена (хотя и намного выше, чем у больных), появляются эритроциты серповидной формы (неполное доминирование А над S). Этот пример показывает, что доминантность может зависеть от условий. Гетерозиготы AS и гомозиготы SS обладают примерно одинаковой устойчивостью к малярии, гомозиготы АА подвержены малярии в большей степени. По данному проявлению ген S доминирует над А. Наконец, в эритроцитах носителей АS в равных количествах присутствуют оба варианта бета-глобиновых цепей — нормальный А и мутантный S (то есть наблюдается кодоминирование).
Неполное доминирование в биологии: что это такое
Благодаря наследственности обеспечивается преемственность поколений. Наследуются не только признаки и свойства организма, но осуществляется передача специфического типа развития.
Взаимоотношение генов и признаков, на которые они оказывают влияние, имеет сложный характер. Один и тот же ген может оказывать влияние на несколько признаков. И в то же время один признак может развиваться под влиянием нескольких генов.
В биологии различают несколько видов доминирования:
- полное;
- неполное;
- кодоминирование;
- аллельное исключение.
Неполным доминированием называется такой тип взаимодействия аллелей, при котором слабый рецессивный признак подавляется сильным доминантным не полностью. При таком явлении для конкретного признака один аллель не доминирует полностью над другим аллелем. В итоге возникает третий фенотип, содержащий признаки доминирующих и рецессивных фенотипов (промежуточный признак).
Такую закономерность называют еще полудоминированием или частичным доминированием.
Первым описал случай неполного доминирования немецкий ботаник Карл Корренс в 1802 году: при скрещивании растений ночной красавицы с красными и белыми цветками он наблюдал появление первого потомства с венчиками розового цвета.
Особенности наследования в случае летальности генов
В некоторых случаях по соотношению фенотипов потомства сложно установить, как взаимодействуют гены. Во втором поколении расщепление при неполном доминировании отличается от ожидаемого 1:2:1, и от 3:1 — при полном. Это бывает, когда доминантный или рецессивный признак дает в гомозиготном состоянии фенотип, не совместимый с жизнью (летальные гены).
У каракульских овец серой окраски новорожденные ягнята, гомозиготные по доминантному аллелю окраски, погибают из-за того, что такой генотип вызывает нарушения развития желудка.
У человека пример летальности доминантной формы гена — брахидактилия (короткопалость). Признак выявляется у в случае гетерозиготного генотипа, в то время как доминантные гомозиготы погибают на ранних этапах внутриутробного развития.
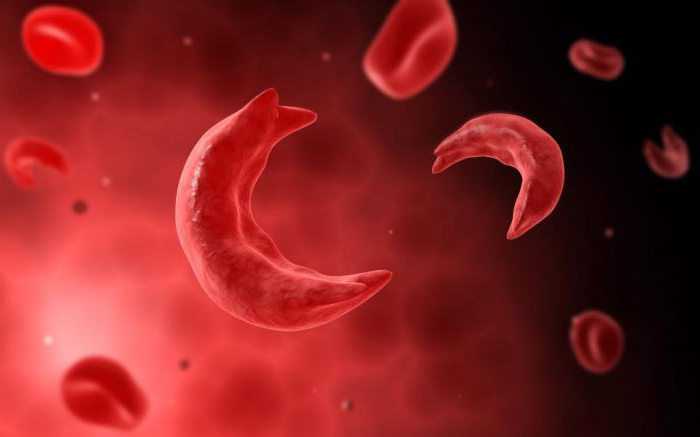
Летальными могут быть и рецессивные аллели генов. Серповидно-клеточная анемия приводит, в случае появления двух рецессивных аллелей в генотипе, к изменению формы эритроцитов. Кровяные клетки не могут эффективно присоединять кислород, и 95% детей с данной аномалией погибают от кислородного голодания. У гетерозигот же измененная форма эритроцитов не влияет в такой степени на жизнеспособность.
Эволюция доминантности [ | ]
Новые мутации могут, конечно, сразу обладать доминантным проявлением в фенотипе диплоидных особей, но вероятность выживания мутантов вообще невелика, и поэтому преимущественно сохраняются именно рецессивные мутации. Впоследствии, если при каких-либо изменениях внешних условий новый признак окажется благоприятным, обусловливающий его мутантный аллель может вторично приобрести доминантное фенотипическое выражение (следует подчеркнуть, что доминантны и рецессивны, собственно говоря, не сами аллели, а их проявления в фенотипе). Переход аллеля от рецессивного к доминантному состоянию может быть обусловлен различными механизмами, действующими на разных уровнях преобразований наследственной информации в онтогенезе. Генетически такой переход может быть достигнут через отбор особых генов-модификаторов, влияющих на фенотипическое проявление мутантного аллеля (гипотеза Р.Фишера), или же через отбор аллелей с большей физиологической активностью (обеспечивающих более интенсивный синтез ферментов), чем первоначальный рецессивный вариант (гипотезы С.Райта и Д.Холдейна). В сущности, эти гипотезы не исключают, а взаимно дополняют друг друга, и эволюция доминантности может происходить путём отбора малых мутаций как структурных генов, так и генов-модификаторов.
Механизмы неполного доминирования
Неполное доминирование – это удивительное генетическое явление это происходит, когда ни один аллель в генная пара является полностью доминантным или рецессивным по отношению к другому. Вместо, уникальный фенотип выражается, когда человек наследует два разных аллеля. Это приводит к сочетанию признаков у потомства, отличающихся от обоих родительских признаков.
Понимание неполного доминирования
Неполное доминирование – это концепция что бросает вызов традиционное понятие of доминирующий и рецессивные аллели. В классическая генетика, доминантные аллели замаскировать выражение рецессивные аллели. Однако в случае неполного доминирования ни один аллель не имеет полный контроль над фенотипом.
Когда человек наследует два разных аллеля специфическая черта, результирующий фенотип is комбинация или смесь черты связанные с каждая аллель. Это смешение наследование приводит к промежуточному фенотипу, отличному от родительские качества.
Пример: Цвет цветка в Snapdragons
Классический пример неполного доминирования можно наблюдать в цвет цветка львиного зева. В этом случае существуют два аллеля участвует: один для красные цветы (R) и еще один для белые цветы (В). Удивительно, но когда растение львиный зев наследуется один красный аллель и один белый аллель (ПВ), получившиеся цветы не красные и не белые, а скорее розовые.
Этот пример ясно демонстрирует, как неполное доминирование может привести к фенотипу, отличному от обоих родительских признаков. Красный аллель не полностью доминирует над аллелем white, и аллель white не является полностью рецессивным. Вместо этого смешивание красные и белые аллели производит уникальный и визуально привлекательный розовый цвет.
Роль генотипа и фенотипа
Чтобы полностью понять неполное доминирование, важно уяснить концепции генотипа и фенотипа. Генотип относится к гентик макияж отдельного человека, в частности сочетание аллелей, которые они наследуют особая черта, На с другой стороны, фенотип наблюдаемая физическая или биохимическая характеристика приводит к взаимодействие между аллелями
В случае неполного доминирования фенотип определяется взаимодействие между два аллеля. Смешение признаков возникает при молекулярный уровень, Где белки Производимые аллели взаимодействуют и влияют на фенотип. Это взаимодействие приводит к промежуточному фенотипу, который комбинация of черты связанные с каждая аллель.
Роль квадратов Пеннета
Квадраты Пеннета – это ценный инструмент в генетике для предсказания генотипы и фенотипы потомства. Их также можно использовать для определения вероятность of разные генотипы и фенотипы в случаях неполного доминирования.
Используя квадраты Пеннета, мы можем визуализировать возможные комбинации аллелей, которые могут быть унаследованы от каждый родитель. В случае неполного доминирования площадь Пеннетта позволяет нам видеть вероятность потомства, наследующего разные аллели и результирующий фенотипs.
Например, в случае львиного зева площадь Пеннета можно использовать для определения вероятность получения розовых цветов при скрещивании львиный зев с красными цветками (RR) с львиный зев с белыми цветками (ВВ). Площадь Пеннетта показывает, что все потомство (RW) будет иметь розовые цветы, так как смешение красные и белые аллели приводит к неполному доминированию.
В заключение следует отметить неполное доминирование. удивительный генетический механизм что бросает вызов традиционное понимание of доминирующий и рецессивные аллели. Это приводит к выражению уникальный фенотип когда человек наследует два разных аллеля. Пример Цвет цветков львиного зева иллюстрирует, как неполное доминирование может привести к появлению фенотипа, отличного от обоих родительских признаков. Понимая роли генотипа и фенотипа, а также используя такие инструменты, как квадраты Пеннета, мы можем получить представление о шаблоны наследования и вероятности, связанные с неполным доминированием.
Новая форма наследования
Иногда в результате скрещивания потомки наследовали промежуточные признаки, которые не давал в гомозиготной форме родительский ген. Неполного доминирования не было в понятийном аппарате генетики до начала XX века, когда были переоткрыты законы Менделя. Тогда же многие естествоиспытатели проводили генетические эксперименты с растительными и животными объектами (томатами, бобовыми, хомяками, мышами, дрозофилами).
После цитологического подтверждения в 1902 г. Уолтером Сеттоном Менделевских закономерностей принципы передачи и взаимодействия признаков стали объяснять с позиции поведения хромосом в клетке.




В том же 1902 году Чермак Корренс описал случай, когда после скрещивания растений с белыми и красными венчиками у потомков были цветки розовой окраски — неполное доминирование. Это проявление у гибридов (генотип Аа
) признака, который является промежуточным по отношению к гомозиготному доминантному (АА
) и рецессивному (аа
) фенотипам. Подобный эффект описан для многих видов цветковых растений: львиного зева, гиацинта, ночной красавицы, ягод земляники.
Взаимодействие аллельных генов
Аллели одного гена, находясь в одном организме, взаимодействуют между собой, и от этого взаимодействия зависит, как проявит себя признак, обусловленный соответствующим геном.
Полное доминирование
Чаще всего встречается такой тип взаимодействия как полное доминирование, при котором один аллельный ген проявляется и полностью подавляет проявление другого аллельного гена. В данном случае первый называется доминантным, а второй — рецессивным.
В генетике принято доминантный ген обозначать большой латинской буквой (например, ), а рецессивный — малой (). Если особь гетерозиготна, то ее генотип будет . Если гомозиготна, то или .
В случае полного доминирования генотипы и будут иметь одинаковый фенотип.
Полное доминирование наблюдается при проявлении, например, цвета семян гороха. У гетерозиготных () растений по этому признаку семена всегда желтые, как и у гомозиготных по доминантному аллелю (). Зеленый цвет определяется рецессивным аллелем и поэтому подавляется.
При множественном аллелизме отношения между аллелями могут быть неоднозначными. Если имеет место быть исключительно полное доминирование, то один ген может быть доминантным по отношению ко второму, но рецессивным по отношению к третьему. В таком случае строят ряды ( > > > …), в которых отражают отношения доминирования. Так, например, наследуется окраска шерсти у многих животных, цвет глаз.
Кроме полного доминирования встречаются и другие типы взаимодействия аллелей: неполное доминирование, кодоминирование, сверхдоминирование, комплементация аллелей и некоторые другие.
Неполное доминирование
В случае неполного доминирования гетерозиготный генотип будет иметь промежуточное значение признака. В этом случае доминантный аллель лишь частично подавляет рецессивный. В результате в фенотипе проявляется некое промежуточное значение признака.
Например, цвет цветков ночной красавицы с генотипом красный, — белый, — розовый.
При неполном доминировании значение признака не обязательно будет ровно «по середине». Чаще бывает отклонение в сторону доминантного, то есть когда фенотип генотипа хоть и не сходен с , но ближе к нему, чем к фенотипу генотипа .
Кодоминирование
При кодоминировании два аллельных гена, оказавшись в одном организме, полностью проявляют себя. Ни один из них никак не подавляет другой. В случае кодоминирования отсутствуют доминантно-рецессивные отношения между аллелями. В итоге в организме синтезируется два разных белка, определяющих один и тот же признак. Нечто похожее происходит при сверхдоминировании и межаллельной комплиментации.
При множественном аллелизме в одной паре аллелей может быть отношение полного доминирования, а в другой — кодоминирования. Так, например, обстоит дело с группами крови человека. Аллель является рецессивным и подавляется доминантными аллелями и . Если организм , то у него первая группа крови. Если или — вторая, или — третья. Если же человек имеет аллели и , то есть генотип , то у него четвертая группа крови, при которой в крови присутствуют два антигена (за наличие одного отвечает аллель , за наличие другого — ).
Сверхдоминирование
Сверхдоминирование — это такой тип взаимодействия аллельных генов, когда у гетерозиготных особей () признак сильнее проявляется, чем у обеих гомозиготных ( и ). Причина подобного явления лежит в достаточно сложных биохимических процессах, связанных с тем, что аллели кодируют несколько разные, но аналогичные по своим функциям полипептиды. В итоге признак как бы накапливается. Сверхдоминирование лежит в основе такого явления как гетерозис, когда дочерние организмы более жизнеспособны, чем родительские.
Помимо перечисленных существуют и другие более редкие типы взаимодействия аллелей.
Неполное доминирование [ | ]
Пример неполного доминирования
При неполном доминировании гетерозиготы имеют фенотип, промежуточный между фенотипами доминантной и рецессивной гомозиготы. Например, при скрещивании чистых линий львиного зева и многих других видов цветковых растений с пурпурными и белыми цветками особи первого поколения имеют розовые цветки. При скрещивании чистых линий андалузских кур чёрной и белой окраски в первом поколении рождаются куры серой окраски. На молекулярном уровне самым простым объяснением неполного доминирования может быть как раз двукратное снижение активности фермента или другого белка (если доминантный аллель дает функциональный белок, а рецессивный — дефектный). Например, за белую окраску может отвечать дефектный аллель, который дает неактивный фермент, а за красную — нормальный аллель, который дает фермент, производящий красный пигмент. При половинной активности этого фермента у гетерозигот количество красного пигмента снижается вдвое, и окраска розовая. Могут существовать и другие механизмы неполного доминирования.
При неполном доминировании во втором поколении моногибридного скрещивания наблюдается одинаковое расщепление по генотипу и фенотипу в соотношении 1:2:1.
В некоторых источниках неполное доминирование характеризуют как такой тип взаимодействия аллелей, когда признак у гибридов F 1 занимает не среднее положение, а отклоняется в сторону родителя с доминирующим признаком. Полностью же средний вариант (как, например, приведённый выше пример наследования окраски цветков) относят к промежуточному характеру наследования
, то есть отсутствию доминирования .