
Возможная цель фотодыхания
Уменьшение фотодыхания может не привести к повышенная скорость роста растений. Фотодыхание может быть необходимо для усвоения нитратов из почвы. Таким образом, снижение фотодыхания с помощью генной инженерии или из-за увеличения содержания углекислого газа в атмосфере (из-за сжигания ископаемого топлива) может не принести пользу растениям, как было предложено. За связь фотодыхания и ассимиляции азота могут быть связаны несколько физиологических процессов. Фотодыхание увеличивает доступность НАДН, который необходим для превращения нитрата в нитрит. Некоторые переносчики нитрита также транспортируют бикарбонат, и было показано, что повышенный уровень CO 2 подавляет транспорт нитрита в хлоропласты. Однако в сельскохозяйственных условиях замена естественного пути фотодыхания на сконструированный синтетический путь метаболизма гликолата в хлоропласте привела к 40-процентному увеличению роста сельскохозяйственных культур.
Хотя фотодыхание значительно снижено в C 4, это все еще важный путь — мутанты без функционирующего метаболизма 2-фосфогликолата не могут расти в нормальных условиях. Было показано, что один мутант быстро накапливает гликолат.
Хотя функции фотодыхания остаются спорными, широко признано, что этот путь влияет на широкий спектр процессов, от биоэнергетики, функции фотосистемы II и метаболизма углерода до ассимиляции азота и дыхание. Оксигеназная реакция RuBisCO может предотвратить истощение CO 2 вблизи его активных центров и способствует регулированию концентрации CO 2. в атмосфере. Фотодыхательный путь является основным источником перекись водорода (H. 2O. 2) в фотосинтетических клетках. Благодаря продукции H. 2O. 2и взаимодействиям пиримидиновых нуклеотидов фотодыхание вносит ключевой вклад в окислительно-восстановительный гомеостаз клеток. Поступая таким образом, он влияет на несколько сигнальных путей, в частности, на те, которые управляют гормональными реакциями растений, контролирующими рост, экологические и защитные реакции, а также запрограммированную гибель клеток.
Было высказано предположение, что фотодыхание может действовать как «безопасность» клапан «, предотвращающий избыточный восстановительный потенциал, возникающий из-за чрезмерно восстановленного НАДФН -пулы, от реакции с кислородом и образования свободных радикалов, поскольку они могут повредить метаболические функции клетки за счет последующего окисления мембранных липидов, белков или нуклеотидов. Мутанты с дефицитом фотодыхательных ферментов характеризуются высоким уровнем окислительно-восстановительного потенциала в клетке, нарушением устьичной регуляции и накоплением формиата.
Полезные аспекты фотодыхания
Сцепление с усвоением нитратов
Если фотодыхание явно ущемляет фотосинтетической активности клеток растительного , было бы полезным и необходимым для других клеточных функций и ограничения не обязательно приведет к лучшему росту растений
Действительно, некоторые исследования показали, что такое фотодыхание может иметь важное значение для фиксации азота в нитратной земле. Считается, что за связь фотодыхания с азотфиксацией ответственны несколько физиологических процессов: с одной стороны, за счет увеличения доступности NADH или NADPH в цитозоле клеток мезофилла , необходимых для восстановления нитратов NO 3 -в нитритах NO 2 -с помощью нитратредуктазы , с другой стороны, за счет ограничения конкуренции бикарбонатов HCO 3 -на носителях из ионов нитрата, эти носители , участвующие в этих двух ионов в то время, и , наконец , позволяя образование достаточного количества ферредоксина приведенных в строму хлоропласта, где он используется в ферредоксин-редуктазы нитрита
С другой стороны, растения с нарушенным метаболизмом 2-фосфогликолата в результате мутации не растут в нормальных условиях, один из этих мутантов показал быстрое накопление гликолата .
Фотодыхание значительно снижено у растений C 4., но, с другой стороны, производит большое количество малата и НАДФН , что сохраняет связывание нитритов.
Редокс-сигнализация
Фотодыхание также может играть роль в передаче сигналов растительными клетками за счет производства перекиси водорода H 2 O 2., молекула, участвующая в передаче сигналов окислительно-восстановительного потенциала, которая контролирует, в частности, механизмы роста и реакции на стресс, например, под действием паразитов.
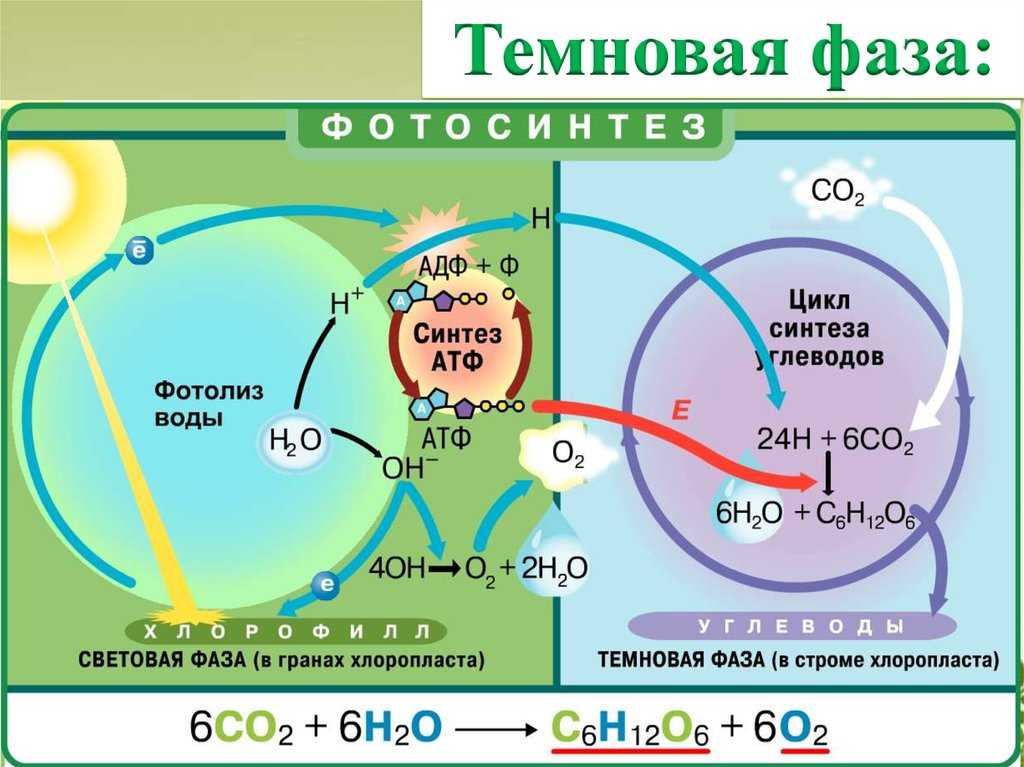
Темновая стадия[]
В темновой стадии с участием АТФ и НАДФН происходит восстановление CO2 до глюкозы. Хотя свет не требуется для осуществления данного процесса, он участвует в его регуляции.
С3 — фотосинтез, цикл Кальвина
Основная статья: Цикл Кальвина
Цикл Кальвина или восстановительный пентозофосфатный цикл состоит из трёх стадий:
- Карбоксилирования
- Восстановления
- Регенерация акцептора CO2
На первой стадии к рибулозо-1,5-бифосфату присоединяется CO2 под действием фермента рибулозобисфосфат-карбоксилаза/оксигеназа (Rubisco). Этот белок составляет основную фракцию белков хлоропласта и предположительно наиболее распространённый фермент в природе. В результате образуется промежуточное неустойчивое соединение, распадающееся на две молекулы 3-фосфоглицериновой кислоты (ФГК).
Во второй стадии ФГК в два этапа восстанавливается. Сначала она фосфорилируется АТФ под действием фосфороглицерокиназы, затем НАДФН при воздействии триозофосфатдегидрогеназы её карбоксильная группа окисляется до альдегидной и она становится углеводом (ФГА).
В третьей стадии участвуют 5 молекул ФГА, которые через образование 4-, 5-, 6- и 7-углеродных соединений объединяются в 3 5-углеродных рибулозо-1,5-бифосфата, для чего необходимы 3АТФ.
Наконец, две ФГА необходимы для синтеза глюкозы. Для образования одной её молекулы требуется 6 оборотов цикла, 6 CO2, 12 НАДФН и 18 АТФ.
С4 — фотосинтез
Основные статьи: Цикл Хетча-Слэка-Карпилова, С4-фотосинтез
При низкой концентрации растворённого в строме CO2 Rubisco катализирует реакцию окисления рибулёзо-1,5-бифосфата и его распад на 3-фосфоглицериновую кислоту и фосфогликолевую кислоту, которая вынужденно используется в процессе фотодыхания.
Для увеличения концентрации CO2 растения С4 типа изменили анатомию листа. Цикл Кальвина у них локализуется в клетках обкладки проводящего пучка, в клетках мезофилла же под действием ФЕП-карбоксилазы фосфоенолпируват карбоксилируется с образованием щавелеуксусной кислоты, которая превращается в малат или аспартат и транспортируется в клетки обкладки, где декарбоксилируется с образованием пирувата, возвращаемого в клетки мезофилла.
С4 фотосинтез практические не сопровождается потерями рибулёзо-1,5-бифосфата из цикла Кальвина, поэтому более эффективен. Однако он требует не 18, а 30 АТФ на синтез 1 молекулы глюкозы. Это оправдывает себя в тропиках, где жаркий климат требует держать устьица закрытыми, что препятствует поступлению CO2 в лист, а также при рудеральной жизненной стратегии.

![Фотодыхание [1983 гэлстон а., дэвис п., сэттер р. - жизнь зеленого растения]](https://lesniepolyani.ru/wp-content/uploads/4/2/a/42a825df2d6dc9a1fd6a5d3e7c54dc4c.png)

САМ фотосинтез
При CAM (Crassulaceae acid metabolism) фотосинтезе происходит разделение ассимиляции CO2 и цикла Кальвина не в пространстве как у С4, а во времени. Ночью в вакуолях клеток по аналогичному вышеописанному механизму при открытых устьицах накапливается малат, днём при закрытых устьицах идёт цикл Кальвина. Этот механизм позволяет максимально экономить воду, однако уступает в эффективности и С4, и С3. Он оправдан при стресстолерантной жизненной стратегии.
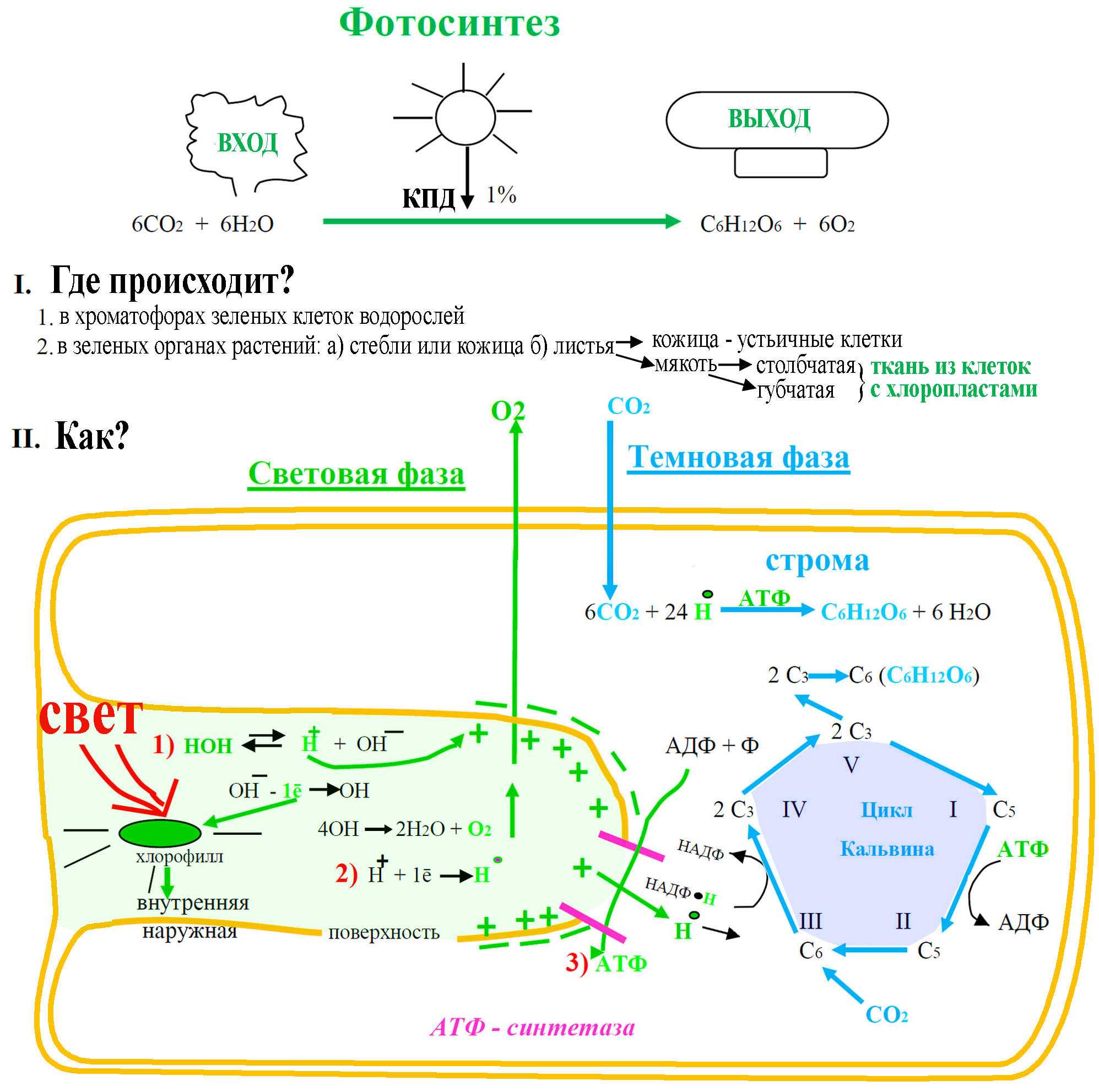
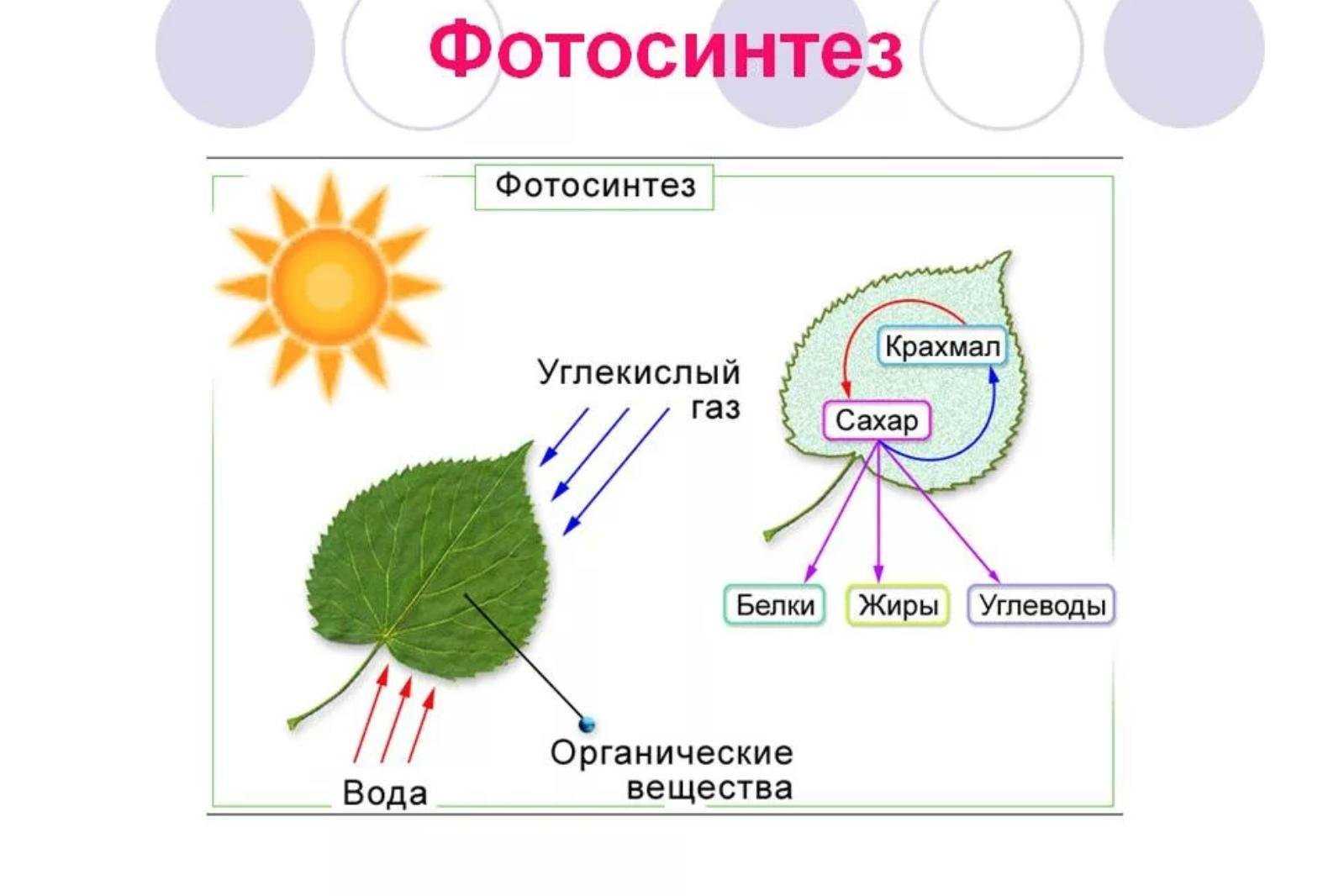
Что такое фотосинтез
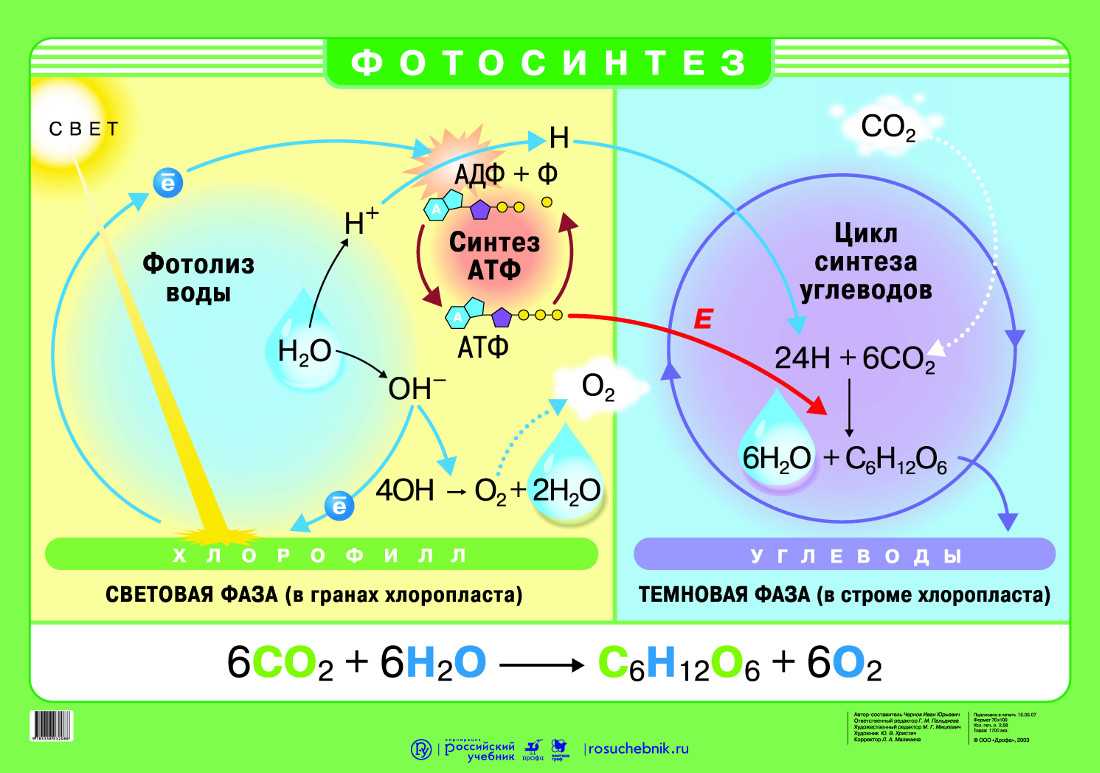
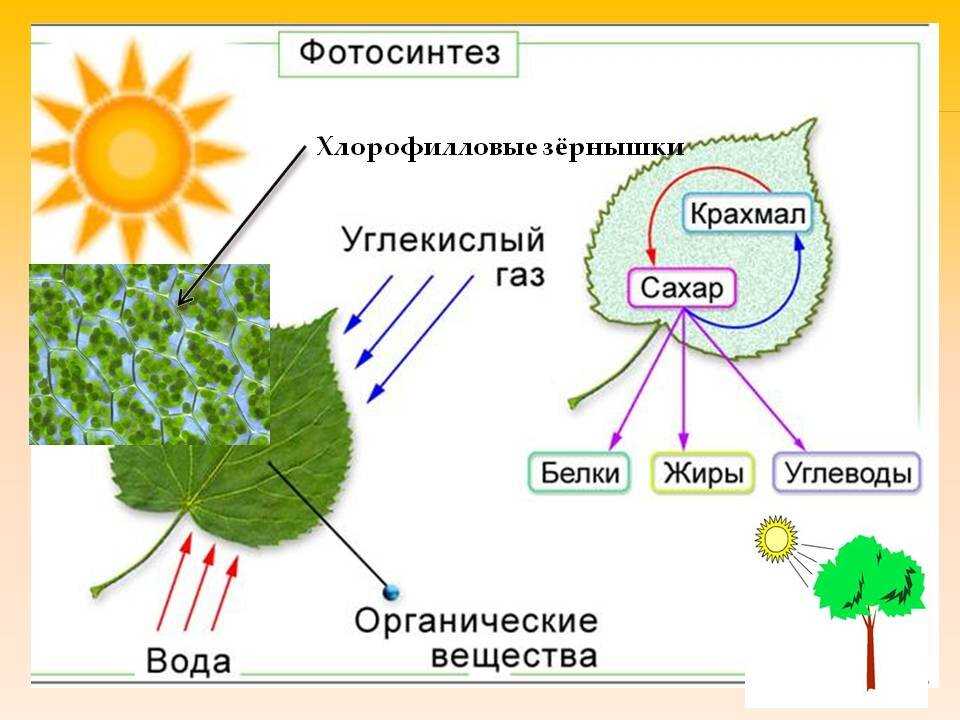
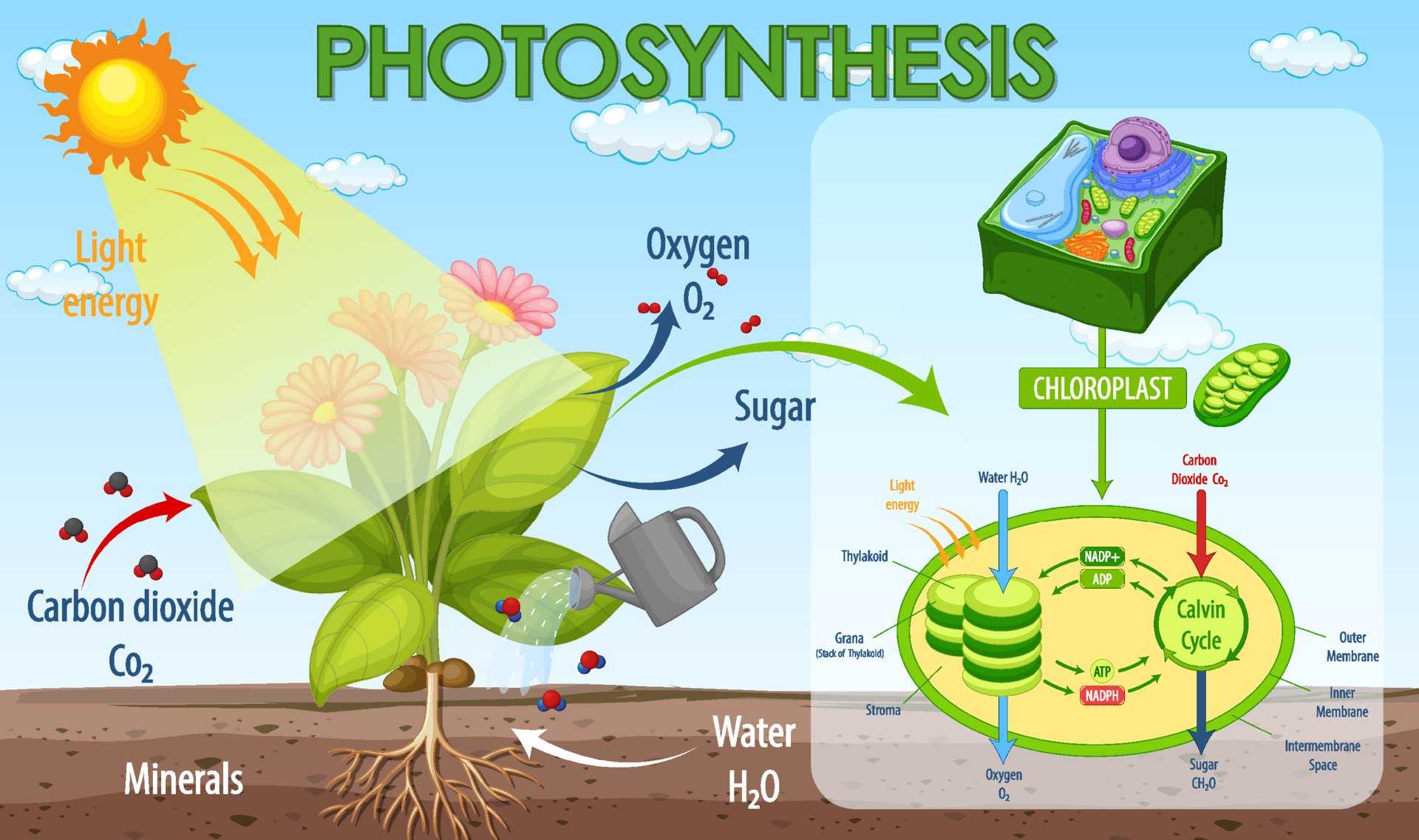
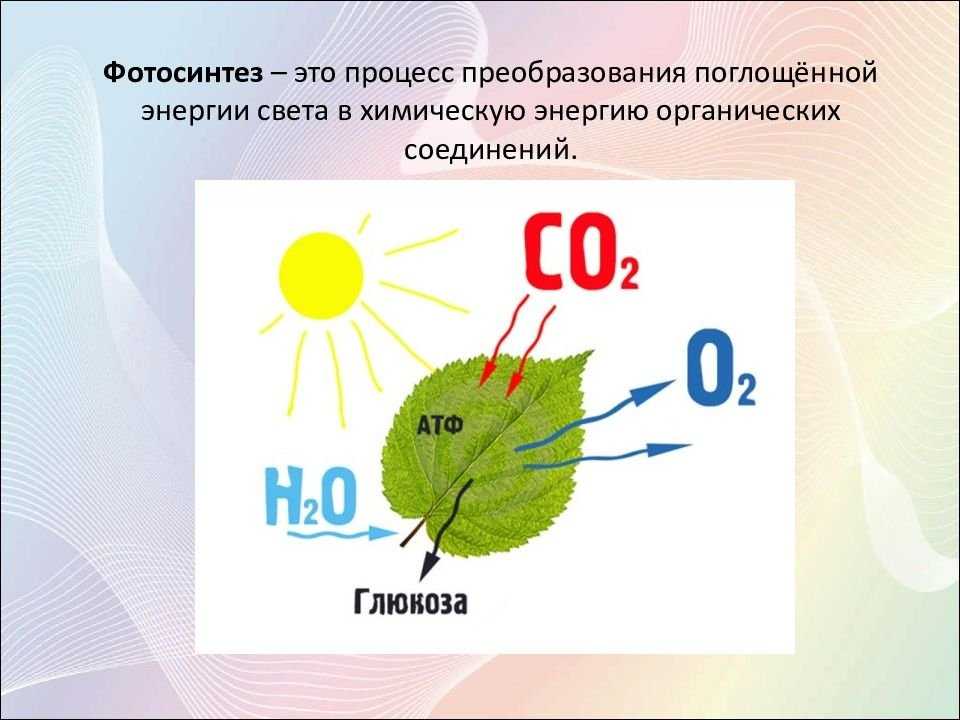
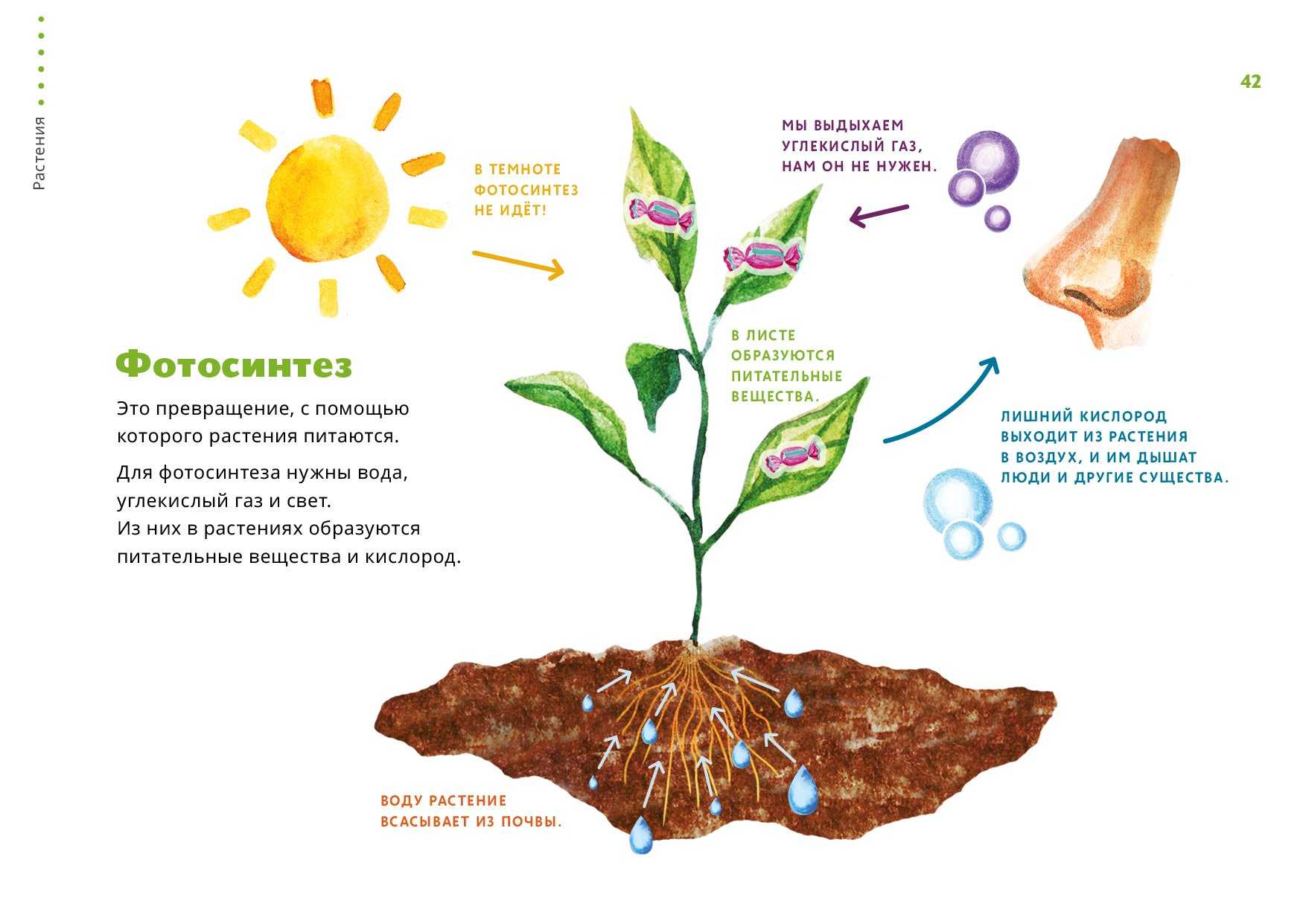
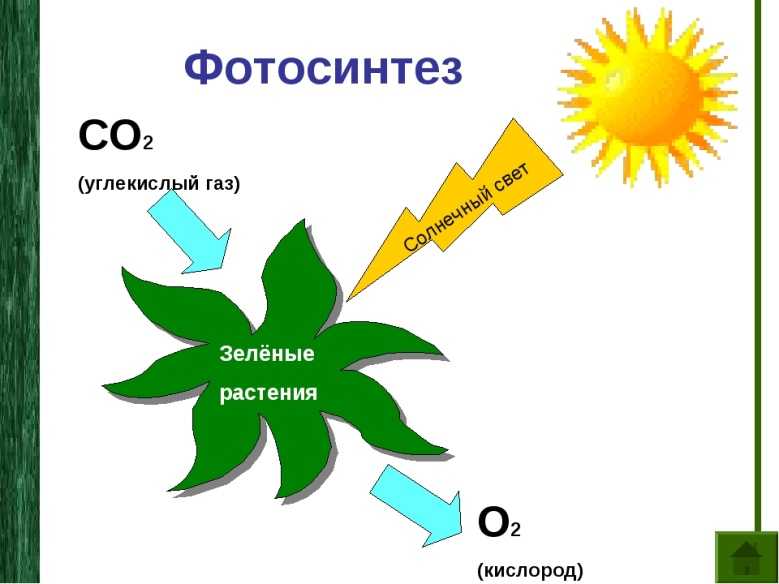
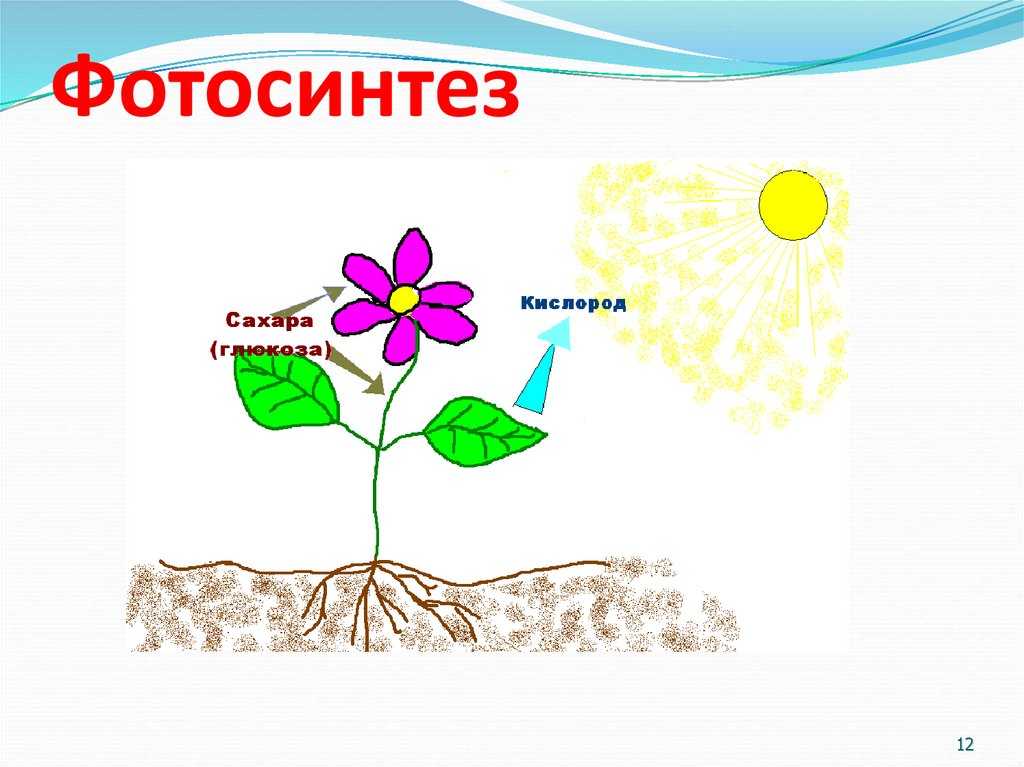
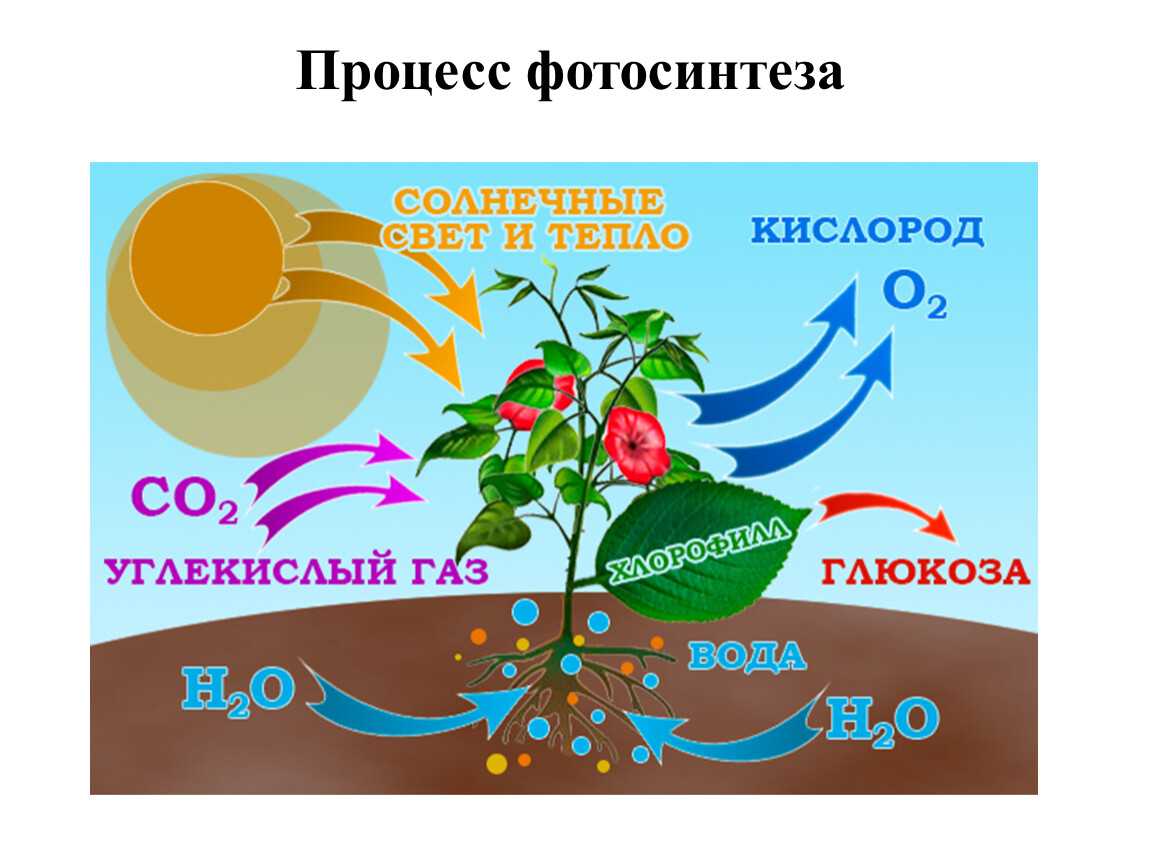
Фотосинтез — это процесс, который производит глюкозу, начиная с углекислого газа и воды, используя энергию солнечного света. Фотосинтетические пигменты, такие как хлорофилл, каротиноиды и фикобилины, улавливают энергию солнечного света. У растений и водорослей эти пигменты концентрируются в хлоропласты. Кислород выделяется как побочный продукт фотосинтеза. Фотосинтез является одним из ключевых процессов, происходящих на Земле, превращающих энергию света в химическую энергию. Глюкоза, полученная в результате этого процесса, может быть использована для производства АТФ в другом процессе, называемом клеточным дыханием.
Процесс фотосинтеза можно разделить на две части: светлая реакция и темная реакция.
Свет реакции
Светлая реакция происходит на тилакоидной мембране граны, стеки тилакоидов внедряются в строму хлоропласта. Фотосинтетические пигменты организованы в фотоцентры на тилакоидной мембране. Фотосистема II поглощает энергию света и переносится в фотоцентры, что позволяет получать электроны высокой энергии. Эти электроны высокой энергии движутся в фотосистему I через комплекс цитохрома b6f. Далее они проходят через ряд носителей ферредоксина, продуцируя НАДФН. Дефицит электронов, возникающий в фотосистемах, восполняется расщеплением молекул воды в процессе, называемом фотолизом. Полученные в результате ионы водорода используются при производстве АТФ.
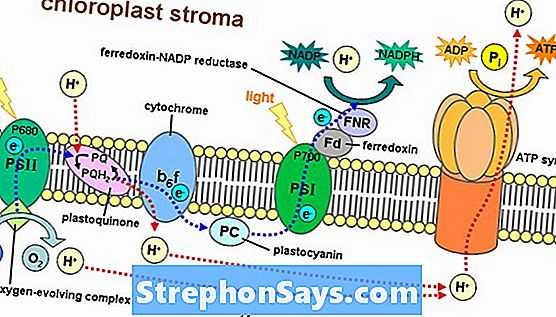
Рисунок 1: Легкая реакция
Темная реакция
Светлая реакция сопровождается темной реакцией. Здесь NADPH и ATP, полученные в результате световой реакции, используются для получения глюкозы из углекислого газа и воды. Темная реакция, которая происходит через цикл С3, также называется циклом Кальвина и происходит в строме хлоропласта без использования света. Фиксация углерода происходит в цикле Кальвина с использованием фермента RuBisCO (рибулозо-1,5-бисфосфаткарбоксилазы / оксигеназы), который фиксирует атом углерода из диоксида углерода в RuBP (рибулозо 1,5-бисфосфат), образуя 3 -phosphoglycerate. Некоторые из молекул 3-фосфоглицерата восстанавливаются, образуя глюкозу, а остальные рециркулируются для производства RuBP. В дополнение к глюкозе, 18 АТФ и 12 NADPH также производятся в течение цикла Кальвина.
Темная реакция, которая происходит в цикле C4, называется путем Хэтча – Слэка, по которому диоксид углерода сначала связывается в PEP, а затем в RuBP.
Взаимодействие органелл при фотодыхании
Осуществление реакций фотодыхания требует тесного взаимодействия трёх органелл растительной клетки: хлоропластов, пероксисом и митохондрий. Выявлен один из интегральных пероксинов PEX10 мембраны пероксисом, обеспечивающий взаимодействие и близкий контакт пероксисомы и хлоропласта. Принципиальным для взаимодействия органелл оказывается C3HC4 (Цис3-Гис-Цис4) RING цинковый палец белка PEX10. Мутация, нарушающая функцию данного домена, оказывается летальной для эмбриона растения на стадии сердца. Рост и развитие у сублетальных мутантов по белку PEX10 нормализуются в условиях обогащенной CO2 среды. В нормальной атмосфере у сублетальных pex10 наблюдается повышенное содержание глиоксилата, сниженное содержание каротиноидов, хлорофиллов a и b, а также сниженный квантовый выход фотосистемы II. При электронной микроскопии наблюдаются изменения в строении пероксисом, а также их разобщенность с хлоропластами. В то же время, точечные мутации в мотиве TLGEEY приводят к формированию нитевидных пероксисом, но при этом не нарушается их ассоциация с хлоропластами. Таким образом, PEX10 контролирует не только ассоциацию пероксисом с хлоропластами, но также их строение.
Примечания
- (недоступная ссылка). Дата обращения: 27 января 2007.
- (недоступная ссылка)
- Zelitch I. // Photosynthesis, Photorespiration, and Plant Productivity (англ.). — New York: Academic Press, 1971. — P. 253—255. — ISBN 0124316085.
- Физиология растений / под ред. И. П. Ермакова. — М. : «Академия», 2007. — 640 с. — ISBN 978-5-7695-36-88-5.
- Фотосинтез. Физиолого-экологические и биохимические аспекты / А.Т Мокроносов, В. Ф. Гавриленко, Т. В. Жигалова; под ред. И. П. Ермакова. — М. : «Академия», 2006. — 448 с. — ISBN 5-7695-2757-9
- ↑
- ↑ Биохимия растений / Г.-В. Хелдт; пер. с англ. — М. : БИНОМ. Лаборатория знаний, 2011. — 471 с. — ISBN 978-5-94774-795-9
Фотодыхание у цианобактерий
В цианобактерии являются единственными бактериями , известными выполняет фотосинтез кислородным. Они достигают фиксации углерода с помощью цикла Кальвина . Долгое время считалось, что они не используют фотодыхание, потому что они концентрируют CO 2.вокруг их Рубиско с помощью специализированных органелл, называемых карбоксисомами , которые сильно усиливают карбоксилазную активность Рубиско в ущерб его оксигеназной активности , и считалось, что образовавшийся небольшой глиоксилат будет легко выведен из клетки без необходимости использования определенного метаболического пути чтобы детоксифицировать его . Тем не менее, мы знаем , что они усваивают на 2-phosphoglycolate в соответствии с набором метаболических путей , которые начинаются прямо на заводе путем преобразования 2-phosphoglycolate в глиоксилате затем GLYCOLATE . Реакции аналогичны реакциям растений для Anabaena и Prochlorococcus marinus , но Synechocystis использует гликолатдегидрогеназу, которая потребляет НАДН и не выделяет перекись водорода H 2 O 2..
Глиоксилат может метаболизироваться по-разному в зависимости от рассматриваемых цианобактерий, и на сегодняшний день частично идентифицированы три метаболических пути:
Он может быть преобразован с помощью глиоксилат — оксидазы в оксалат , сама преобразуется в оксалатоксидазы и формиатдегидрогеназы на две молекулы из CO 2.
Он также может быть преобразован в гидроксипируват , как в растениях, или путем конденсации двух молекул глиоксилата с помощью тартронат-полуальдегидсинтазы с образованием полуальдегида тартроната , который сам превращается в глицерат с помощью полуальдегидредуктазы тартроната с потреблением одной молекулы NADH или NADPH .
В Synechocystis и Anabaena , то hydroxypyruvate снижается глицерат претерпевает фосфорилирование с помощью 3-фосфоглицерата под действием глицерата киназы , как в растениях, но и другие цианобактерии первой форма 2-фосфоглицерата , изомеризуется затем к 3-фосфоглицерату посредством реакции , аналогичной катализируются фосфоглицератмутаз из гликолиза .
Эти три метаболических пути пересекаются и происходят одновременно у Synechocystis . Только когда все три инактивированы, этой цианобактерии, как и растениям, требуется высокая концентрация углекислого газа, чтобы выжить.
Относительное сродство Рубиско к кислороду и углекислому газу
Активный сайт из Рубиско имеет особенность , имеющие как карбоксилазы активности и оксигеназу активность . Это связано с тем, что фермент активирует свой субстрат , рибулозо-1,5-бисфосфат , как нестабильный эндиол, который также может реагировать с CO 2.чем с O 2 :
Рибулозо-1,5-бисфосфат ( 1 ) активируется Рубиско как enediol ( 2 ) , способного реагировать как с CO 2дать две молекулы 3-фосфоглицерата ( 3 ) с O 2с образованием молекулы 3-фосфоглицерата ( 3 ) и молекулы 2-фосфогликолата ( 4 ).
Тот факт , что насыщение кислородом происходит примерно в 25% случаев по сравнению с карбоксилирования предполагает , что последний сильно благоприятствует Рубиско , если принять во внимание тот факт , что кислород находится примерно в 500 раз более обильными в атмосфере. В двуокись углерода , это отношение упало 25 раз в растворе в воде до 25 ° C
Коэффициент селективности по CO 2/ O 2колеблется от 20 до 280 в зависимости от вида , а скорость катализа колеблется от 2 до 8 циклов в секунду.
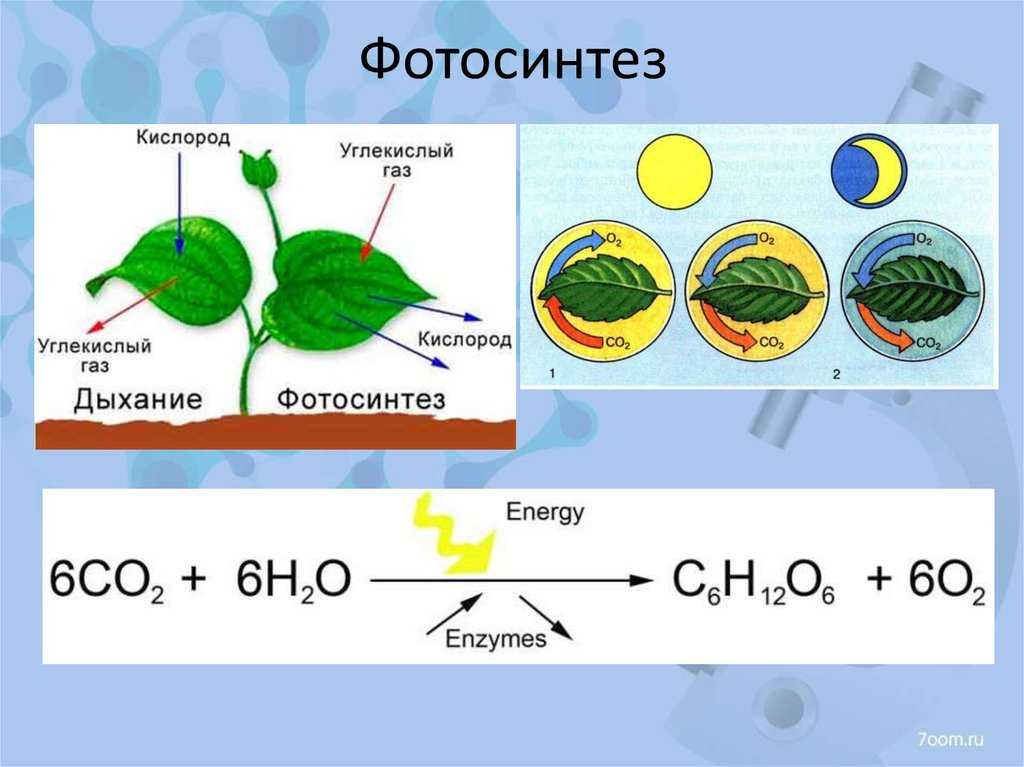
Газообмен и отличия от темнового дыхания
Суммарное поглощение кислорода на свету определяется интенсивностью двух процессов: темновым дыханием и светоиндуцируемым дыханием, связанным с фотохимическими реакциями хлоропластов. Фотодыхание активируется при высоких интенсивностях света, в то время как процессы темнового дыхания растений подавляются на свету. Фотодыхание увеличивается при повышении концентрации кислорода от 0 до 100 %, а темновое дыхание насыщается уже при 2 % кислорода. В отличие от темнового дыхания процесс поглощения кислорода при фотодыхании не ингибируется типичными дыхательными ядами, например азидом натрия (ингибитор цитохромоксидазы митохондрий).

Биологическое значение фотодыхания
Хотя достоверно известно, что фотодыхание снижает эффективность фотосинтеза, приводит к потерям ассимилированного углерода, тем не менее вопрос о функциях фотодыхания остается дискуссионным. Основная гипотеза предполагает, что фотодыхание возникло как путь, служащий для оптимальной утилизации гликолата, образующегося в результате оксигеназной активности РуБисКО. При этом тот факт, что оксигеназная активность РуБисКО не была элиминирована в ходе эволюции, объясняется, по-видимому тем, что существующее соотношение карбоксилазной и оксигеназной активностей достигло предела, определяемого химизмом реакции и не может быть увеличено. Сравнительный анализ фермента разных организмов показывает, что РуБисКО существовала уже 3,5 млрд лет назад, когда в атмосфере было мало кислорода и уже к тому времени заняла ключевую позицию в цикле ассимиляции углерода при фотосинтезе. При этом её оксигеназная функция в условиях низкого содержания кислорода не играла существенной роли. По мере увеличения содержания кислорода потери ассимилированного углерода в результате фотодыхания нарастали, однако сложность строения РуБисКО, по-видимому, помешала эволюции каталитического центра устранить оксигеназную активность. Данная гипотеза косвенно подтверждается отсутствием значительных успехов в попытках генноинженерным способом увеличить сродство РуБисКО к углекислому газу путём изменения аминокислотной последовательности активного центра фермента. Фотодыхание, вследствие оксигеназной реакции РуБисКО, предотвращает исчерпание углекислого газа у активного центра этого фермента и, в конечном итоге, регулирует содержание CO2 и O2 в биосфере.
В связи с тем, что фотодыхание снижает эффективность фотосинтеза, в ходе эволюции у ряда растений возникли минимизирующие фотодыхание механизмы, не связанные с модификацией РуБисКО. К таким механизмам относятся различные типы C4-фотосинтеза и CAM-фотосинтез. В этих биохимических путях первичную фиксацию углекислоты осуществляет фосфоенолпируваткарбоксилаза (ФЕП-карбоксилаза), что позволяет в конечном счёте концентрировать углекислоту в месте её ассимиляции в реакции карбоксилирования РуБФ, катализируемой РуБисКО.
Эволюция
Предшественники современных цианобактерий были первыми организмами, способными к кислородному фотосинтезу . Таким образом, они также являются первыми организмами, которым Рубиско подвергся воздействию кислорода . Они, безусловно, должны были произвести 2-фосфогликолят, потому что механизмы концентрации CO 2появился намного позже, вероятно, примерно за 360–300 миллионов лет до настоящего времени, когда концентрация кислорода в атмосфере значительно увеличилась. С тех пор растения утратили некоторые ферменты метаболизма глицерата , которые все еще присутствуют в цианобактериях, но фотодыхание никуда не делось. Даже бактериопланктон , такой как Prochlorococcus marinus и Synechococcus , которые имеют очень маленький геном , сохранили гены фотодыхания. Этот метаболический путь либо присутствовал с самого начала у цианобактерий, либо появился очень рано у предшественников последних.
Согласно симбиогенезу , предшественники нынешних цианобактерий также являются предшественниками хлоропластов этих клеток из растений и водорослей . Рубиско всех этих организмов (цианобактерии, растения, водоросли) объединяет то, что они также принимают кислород O 2.чем углекислый газ CO 2реагировать с рибулозо-1,5-бисфосфатом . Селективность Рубиско по CO 2относительно O 2 вероятно, не сильно увеличился в процессе эволюции, потому что этот фермент по существу эволюционировал бы, пока концентрация кислорода в атмосфере была все еще низкой, так что фотодыхание всегда оставалось незначительным и не могло быть фактором, определяющим отбор.
§ 22. Фотосинтетические пигменты. Световая фаза фотосинтеза
В отличие от гетеротрофов, которым необходимы готовые органические соединения, автотрофы способны синтезировать их из неорганических веществ. Процессы биосинтеза относятся к пластическому обмену и, следовательно, сопровождаются поглощением энергии. Из курса биологии 10-го класса вам известно, что некоторые бактерии (железобактерии, бесцветные серобактерии, нитрифицирующие, водородные) для образования органических соединений используют энергию, которая выделяется при окислении неорганических веществ. Такие организмы называются хемоавтотрофами. Однако большинство автотрофных организмов использует для этого энергию света. Они составляют группу фотоавтотрофов.
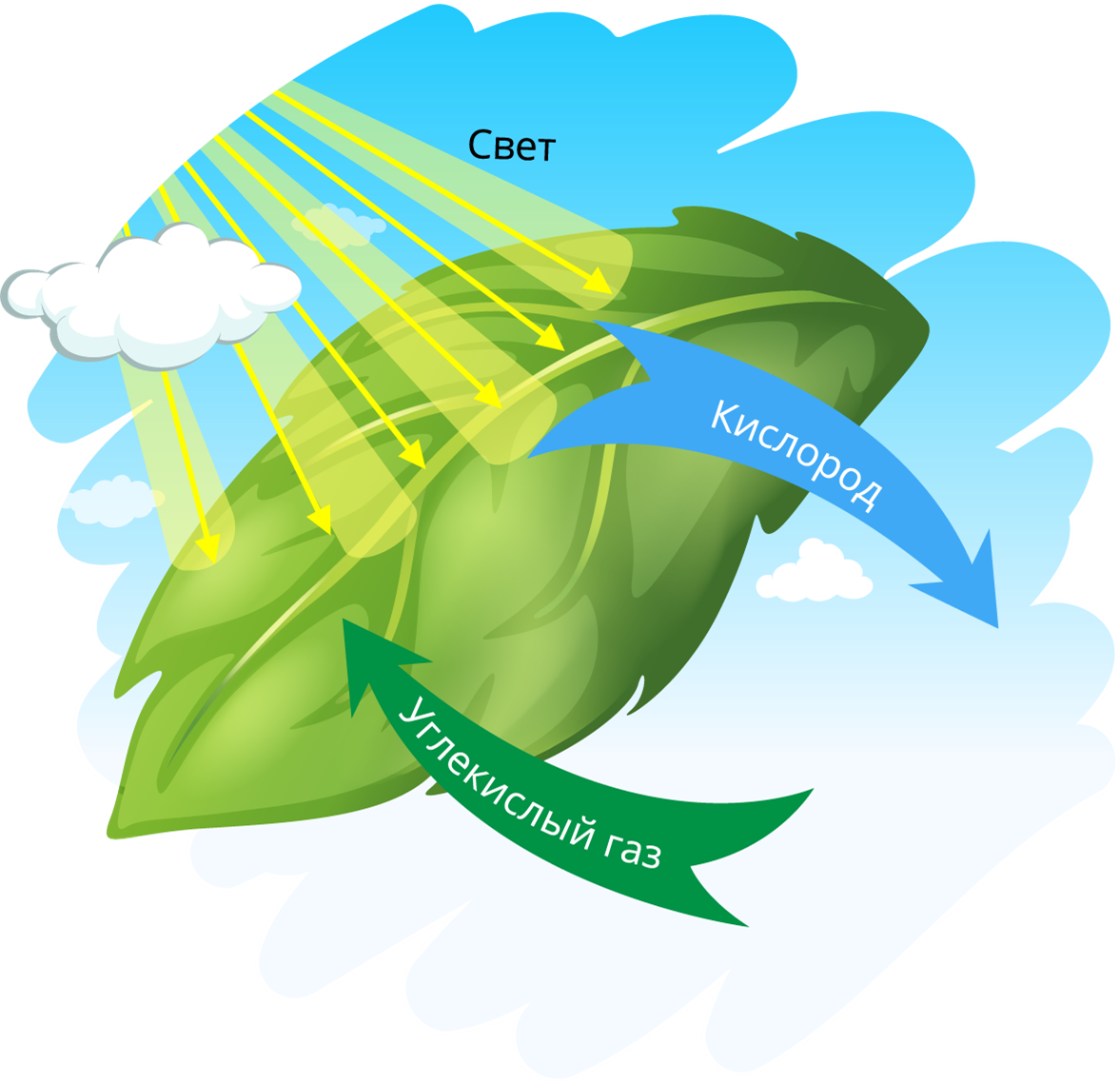
Понятие фотосинтеза. К фотоавтотрофам относятся зеленые растения, водоросли, цианобактерии *зеленые и пурпурные серобактерии* и др. В процессе фотосинтеза с помощью специальных пигментов они поглощают световую энергию и преобразуют ее в энергию химических связей органических веществ. Исходным материалом для синтеза органических соединений являются такие неорганические вещества, как углекислый газ и вода. Таким образом, фотосинтез — это процесс образования органических веществ из углекислого газа и воды, протекающий с поглощением энергии света при участии фотосинтетических пигментов.
*В XVII в. нидерландский исследователь Я. ван Гельмонт посадил в горшок с землей ветвь ивы, предварительно взвесив как почву, так и побег растения. В течение 5 лет он выращивал иву, не добавляя в горшок ничего, кроме воды. Затем ученый снова взвесил почву и растение. Оказалось, что масса ивы за эти годы увеличилась почти на 75 кг, в то время как масса почвы уменьшилась всего на 57 г. На основании этого ван Гельмонт пришел к выводу о том, что вещества растений образуются только из воды, а не из почвы и воздуха. Это ошибочное мнение было распространено до конца XVIII в.
В 1771 г. британский естествоиспытатель Дж. Пристли поместил живую веточку мяты в закрытый сосуд, воздух в котором был «испорчен горением свечи». Через несколько дней он обнаружил, что свеча в этом сосуде снова могла гореть, и сделал заключение о том, что растения способны «исправлять воздух». Следовательно, открытие фотосинтеза принадлежит Дж. Пристли. В то время он полагал, что воздух — это единое вещество, которое переходит из одной формы в другую. Дальнейшие опыты привели Дж. Пристли к открытию кислорода, и в 1778 г. он доказал, что при фотосинтезе растения поглощают углекислый газ и выделяют кислород. Когда Дж. Пристли награждали медалью за его достижения, ученый сказал: «Благодаря этим открытиям мы уверены, что растения произрастают не напрасно, а очищают и облагораживают нашу атмосферу».
Однако Дж. Пристли не обратил внимания на то, что фотосинтез протекает только при наличии света. Это установил голландский физиолог Я. Ингенхауз. Он погружал ветвь растения в воду и наблюдал выделение пузырьков кислорода на свету. В темноте образование кислорода прекращалось. В 1796 г. Ингенхауз вывел следующее уравнение фотосинтеза:
Пространственная локализация[]
Хлоропласты в клетках листа
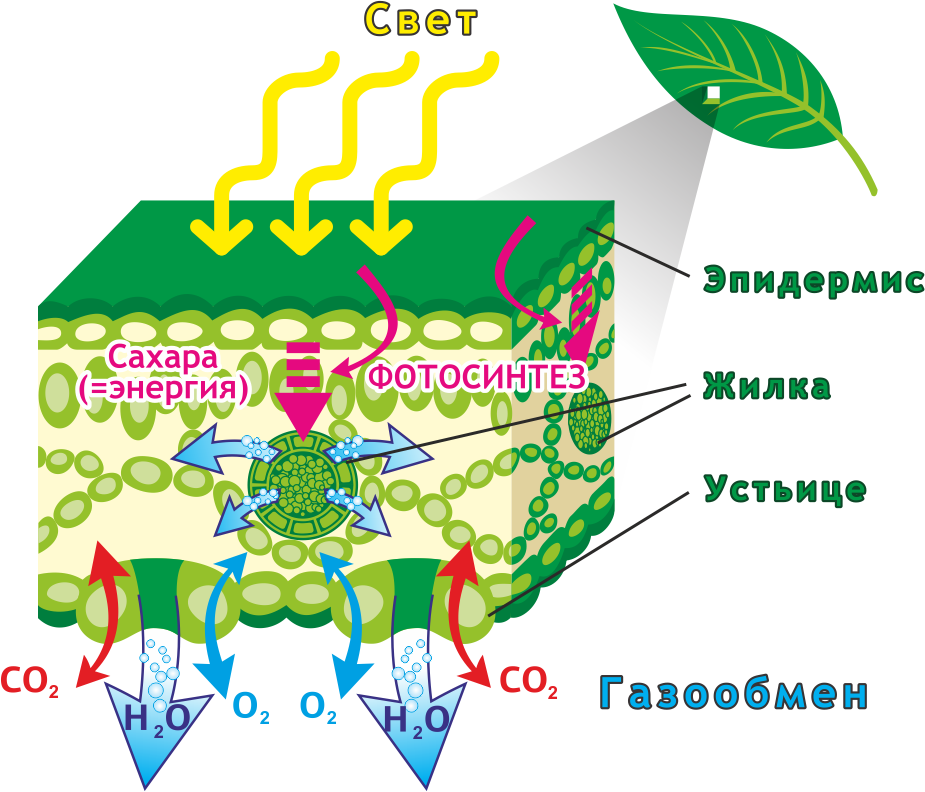
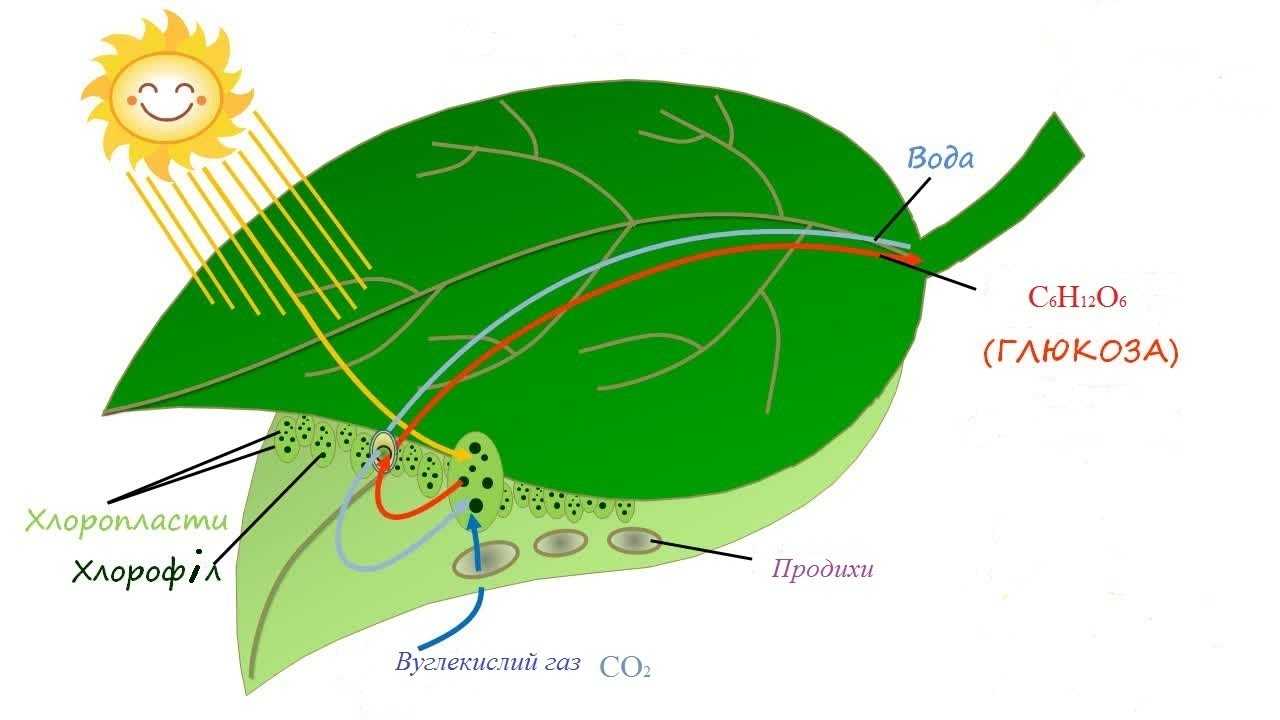
Фотосинтез растений осуществляется в хлоропластах, обособленных двухмембранных органеллах клетки. Хлоропласты могут быть в клетках плодов, стеблей, однако основным органом фотосинтеза, анатомически приспособленным к его ведению, является лист. В листе наиболее богата хлоропластами ткань палисадной паренхимы. У некоторых суккулентов с вырожденными листьями (например, кактусы) основная фотосинтетическая активность связана со стеблем.
Свет для фотосинтеза захватывается более полно благодаря плоской форме листа, обеспечивающей большое отношение поверхности к объёму. Вода доставляется из корня по развитой сети сосудов (жилок листа). Углекислый газ поступает отчасти посредством диффузии через кутикулу и эпидермис, однако большая его часть диффундирует в лист через устьица и по листу по межклеточному пространству. Растения, осуществляющие С4 и CAM фотосинтез сформировали особые механизмы для активной ассимиляции углекислого газа.
Внутреннее пространство хлоропласта заполнено бесцветным содержимым (стромой) и пронизано мембранами (ламеллами), которые соединяясь друг с другом образуют тилакоиды, которые в свою очередь группируются в стопки, называемые граны. Внутритилакоидное пространство отделено и не сообщается с остальной стромой, предполагается также что внутреннее пространство всех тилакоидов сообщается между собой. Световые стадии фотосинтеза приурочены к мембранам, автотрофная фиксация CO2 происходит в строме.
В хлоропластах имеются свои ДНК, РНК, рибосомы (70s типа), идёт синтез белка (хотя этот процесс и контролируется из ядра). Они не синтезируются вновь, а образуются путём деления предшествующих. Всё это позволило считать их предшественниками свободные цианобактерии, вошедшие в состав эукариотической клетки в процессе симбиогенеза.
Цианобактерии, таким образом, как бы сами являются хлоропластом и в их клетке фотосинтетический аппарат не вынесен в особую органеллу. Их тилакоиды, однако, не образуют стопок, а формируют различные складчатые структуры (у единственной цианобактерии Gloeobacter violaceus тилакоиды отсутствуют вовсе, а весь фотосинтетический аппарат находится в цитоплазматической мембране, не образующей впячиваний). У них и растений также есть различия в светособирающем комплексе (см. ниже) и пигментном составе.
Субстратная специфичность RuBisCO
Окислительная реакция фотосинтетического углеродного цикла катализированный к RuBP оксигеназа Мероприятия:
- RuBP + О2 → Фосфогликолят + 3-фосфоглицерат + 2ЧАС+
Оксигеназная активность RuBisCO
Во время катализа RuBisCO в активном центре RuBisCO образуется «активированный» интермедиат (эндиоловый интермедиат). Это промежуточное соединение способно реагировать либо с CO2 или же О2. Было продемонстрировано, что особая форма активного сайта RuBisCO стимулирует реакции с CO2. Хотя существует значительный процент «неудач» (~ 25% реакций представляют собой оксигенацию, а не карбоксилирование), это представляет собой значительное преимущество CO2, когда учитывается относительное содержание двух газов: в текущей атмосфере, О2 примерно в 500 раз больше, а в растворе О2 в 25 раз больше, чем CO2.
Способность RuBisCO определять между двумя газами известна как его коэффициент селективности (или Srel), и он варьируется между видами, с покрытосеменными более эффективными, чем другие растения, но с небольшими вариациями среди сосудистые растения.
Предлагаемое объяснение неспособности RuBisCO полностью различать CO2 и О2 в том, что это эволюционный реликт:[нужна цитата ] Ранняя атмосфера, в которой возникли примитивные растения, содержала очень мало кислорода, ранняя эволюция RuBisCO не находился под влиянием его способности различать О2 и CO2.
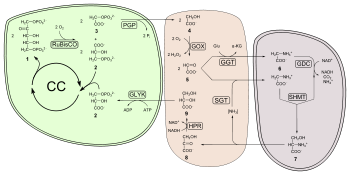
Фотодыхательные реакции
 хлоропластпероксисомамитохондрия
хлоропластпероксисомамитохондрия
Добавление молекулярного кислорода к рибулозе-1, 5-бисфосфат дает 3-фосфоглицерат (PGA) и 2-фосфогликолят (2PG или PG). PGA является нормальным продуктом карбоксилирования и продуктивно входит в цикл Кальвина. Однако фосфогликолят подавляет определенные ферменты, участвующие в фотосинтетической фиксации углерода (поэтому его часто называют «ингибитором фотосинтеза»). Его также относительно сложно перерабатывать: у высших растений он восстанавливается с помощью серии реакций в пероксисоме, митохондриях и снова в пероксисоме, где он превращается в глицерат. Глицерат повторно поступает в хлоропласт тем же транспортером, который экспортирует гликолят. Стоимость 1 АТФ связана с преобразованием в 3-фосфоглицерат (PGA) (Фосфорилирование ) в хлоропласте, который затем может повторно поступать цикл Кальвина.


![Фотодыхание [1983 гэлстон а., дэвис п., сэттер р. - жизнь зеленого растения]](https://lesniepolyani.ru/wp-content/uploads/b/d/0/bd07b329e7165015469689c81b0e1f36.png)
С этим метаболическим путем связано несколько затрат; образование пероксида водорода в пероксисоме (связанное с превращением гликолата в глиоксилат). Перекись водорода является опасно сильным окислителем, который должен немедленно расщепляться на воду и кислород ферментом каталазой. Превращение 2 × 2Carbon глицина в 1 C3 серин в митохондриях ферментом глицин-декарбоксилазой является ключевым этапом, который высвобождает CO. 2, NH 3., и восстанавливает НАД до НАДН. Таким образом, 1 молекула CO. 2образуется на каждые 2 молекулы O. 2(две происходят от RuBisCO, а одна — от пероксисомального окисления). Ассимиляция NH 3 происходит через цикл GS -GOGAT за один АТФ и один НАДФН.
Цианобактерии могут метаболизировать 2-фосфогликолят тремя способами. Они не могут расти, если нарушены все три пути, несмотря на наличие механизма концентрирования углерода, который должен резко снизить скорость фотодыхания (см. Ниже).
Рекомендации
-
(in) Томас Д. Шарки , » Оценка скорости фотодыхания в листьях « , Physiologia Plantarum , vol. 73, п о 1,
Май 1998 г., стр. 147-152 -
(in) Ричард Ли Гуд , Желанное отвлечение от фотодыхания , Nature Biotechnology , Vol. 25,
2007 г., стр. 539-540 -
↑ и (in) Герман Бауве, Мартин Хагеманн и Алисдэр Р. Ферни , Фотодыхание: участники, партнеры и происхождение , Trends in Plant Science , vol. 15, п о 6,
июнь 2010, стр. 330-336 -
(in) Марион Эйзенхут, Вольфганг Рут Майя Хаймович, Герман Бауве Аарон Каплан и Мартин Хагеманн , Фотодыхательный метаболизм гликолата необходим для цианобактерий и, возможно, был передан эндосимбионтным растениям , PNAS , vol. 105, п о 44,
4 ноября 2008 г., стр. 17199-17204 -
(in) Вероника Дж. Маурино и Кристоф Петерхансель , Фотодыхание: современное состояние и подходы к метаболической инженерии , Current Opinion in Plant Biology , vol. 13, п о 3,
июнь 2010, стр. 248-255 -
(in) Говард Гриффитс , Биология растений: Дизайн Рубиско , Nature , vol. 441, г.
22 июня 2006 г., стр. 940-941 -
(ru) Шимон Рахмилевич Асаф Б. Казинс и Арнольд Дж. Блум , Ассимиляция нитратов в побегах растений зависит от фотодыхания , PNAS , vol. 101, п о 31,
3 августа 2004 г., стр. 11506-11510 -
(in) Арнольд Дж. Блум, Мартин Бургер, Хосе Сальвадор Рубио Асенсио Асаф Б. Казинс , Обогащение углекислым газом препятствует ассимиляции нитратов в пшенице и арабидопсисе , Science , vol. 328, п о 5980,
14 мая 2010 г., стр. 899-903 -
(in) Эдуардо Забалета, Виктория М. Мартин и Ханс-Петер Браун , Механизм концентрации основного углерода в растениях? , Растениеводство , т. 187, г.
Май 2012 г., стр. 97-104 -
(in) Кристин Х. Фойер, Арнольд Дж. Блум, Уильям Грэм и Кувал Ноктор , Фотодыхательный метаболизм: гены, мутанты, энергия и редокс-сигналы , Annual Review of Plant Biology , vol. 60,
Июнь 2009 г., стр. 455-484 -
(in) Ингер Андерссон , » Катализ и регулирование в Рубиско « , Журнал экспериментальной ботаники , Vol. 59, п о 7,
2008 г., стр. 1555-1568 -
(in) Сигрун Рейманн PM и Андреас Вебер , Пероксисомы растений дышат светом: некоторые пробелы фотодыхания C 2
цикл наполнился — другие остались » , Biochimica et Biophysica Acta (BBA) — Molecular Cell Research , vol. 1763, п о 12,декабрь 2006 г., стр. 1496-1510 - (in) Дональд Р. Орт , Хелен В. Лю , Аманда П. Кавана и Пол Ф. Саут , « Пути метаболизма синтетического гликолята стимулируют рост сельскохозяйственных культур и урожайность в поле » , Science , vol. 363, п о 6422,4 января 2019 г.,, eaat9077
Условия, влияющие на фотодыхание
Показатели фотодыхания увеличиваются:
Измененная доступность субстрата: пониженный уровень CO2 или повышенный O2
Факторы, которые влияют на это, включают в себя содержание двух газов в атмосфере, подачу газов к месту фиксации (то есть на наземных растениях: наличие устьица открыты или закрыты), длина жидкой фазы (насколько далеко эти газы должны диффундировать через воду, чтобы достичь места реакции). Например, когда устьица закрыты для предотвращения потери воды во время засуха: это ограничивает CO2 поставка, в то время как О2 производство внутри листа будет продолжено. В водорослях (и растениях, фотосинтезирующих под водой); газы должны распространяться через воду на значительные расстояния, что приводит к снижению доступности CO2 относительно О2. Было предсказано, что увеличение окружающий CO2 концентрации, прогнозируемые в течение следующих 100 лет, могут снизить скорость фотодыхания у большинство растений примерно на 50%[нужна цитата ]. Однако при температурах выше, чем термальный оптимум фотосинтеза, увеличение скорости обмена не приводит к усилению ассимиляции СО2 из-за пониженного сродства Рубиско к СО2.
Повышенная температура
При более высоких температурах RuBisCO менее способен различать CO2 и О2. Это потому, что промежуточное соединение эндиола менее стабильно. Повышение температуры также снижает растворимость CO2, тем самым снижая концентрацию CO2 относительно О2 в хлоропласт.