

Полимерное взаимодействие доминантных неаллельных генов
Явление одновременного действия на признак нескольких неаллельных однотипных генов получило название полимерии. Хотя полимерные гены не являются аллельными, но так как они определяют развитие одного признака, их обычно обозначают одной буквой А (а), цифрами указывая число аллельных пар.
Действие полигенов чаще всего бывает суммирующим. Признаки, зависящие от полимерных генов, относят к количественным признакам. В качестве примера можно привести наследование окраски зерен у пшеницы. Полигенное взаимодействие генов открыто шведским ученым Нильсоном-Эле.
Задача 8
На модификацию дигибридного скрещивания при полимерии в отношении 15:1
| Ген | Признак |
| А1 А2 A1a2 | окрашенные зерна окрашенные зерна белые зерна |
Фенотипы окраш. зерна х белые зерна
Родителей
Во втором поколении наблюдается расщепление в отношении 15:1; 1/16 а1а1а2а2зерен пшеницы не имеют окраски. Наиболее окрашенными являются растения с генотипами А1А1А2А2 — 1/16. Чем больше доминантных генов в генотипе, тем интенсивнее окраска.
У человека примером полимерии является наследование пигментации кожных покровов.
Генетический контроль количества образуемого меланина осуществляется не зависимо четырьмя полимерными генами: P1P2P3P4- При наличии доминантных аллелей этих генов синтезируется много пигментов.
Она максимальна у африканцев, которые являются — доминантными тетрагомозиготами Р1Р1Р2Р2,Р3Р4Р4, и минимальная у европеоидов — рецессивных тетрагомозигот по соответствующим локусам р1р1,р2р2,р3р3,р4р4. В браках африканцев и европейцев рождаются мулаты тетрагетерозиготы, имеющие по сравнению с родителями промежуточные значения интенсивности пигментации кожных покровов: Р1РьР2Р2,Рзрз,Р4Р4.В браках между мулатами рождаются дети с пигментацией кожи от максимально темной до максимально светлой.
Важная особенность полимерии суммирование (аддитивность) или кумулятивность действия неаллельных доминантных генов на развитие количественных признаков. Многие морфологические признаки человека, а также патологические особенности, определяются полимерными генами: рост, масса тела, величина артериального давления и т.д.
Модификация дигибридного скрещивания при полимерном взаимодействии генов 15:1; тригибридного 63:1; тетрагибридного 255:1
Плейотропия — явление, когда один и тот же ген может действовать на различные признаки организма. Однако не следует представлять, что плейотропный ген в равной степени влияет на каждый из признаков. Для абсолютного большинства генов с той или иной степенью плейотропии характерно сильное влияние на один признак и значительно более слабое влияние на другие. Так, ген, определяющий рыжую окраску волос, одновременно обусловливает более светлую окраску кожи и появление веснушек.
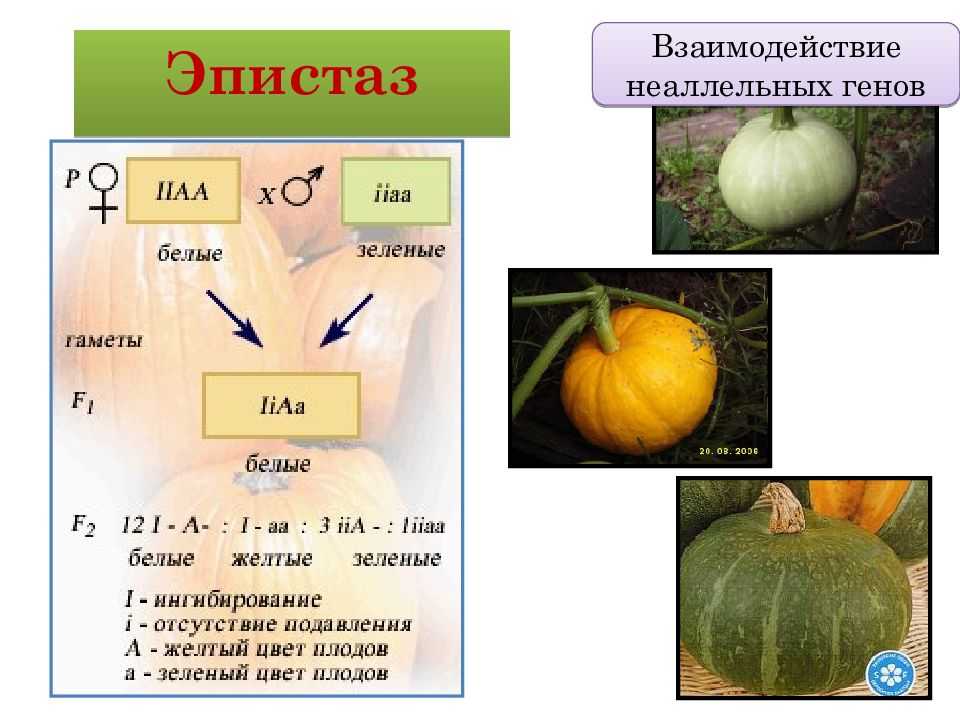
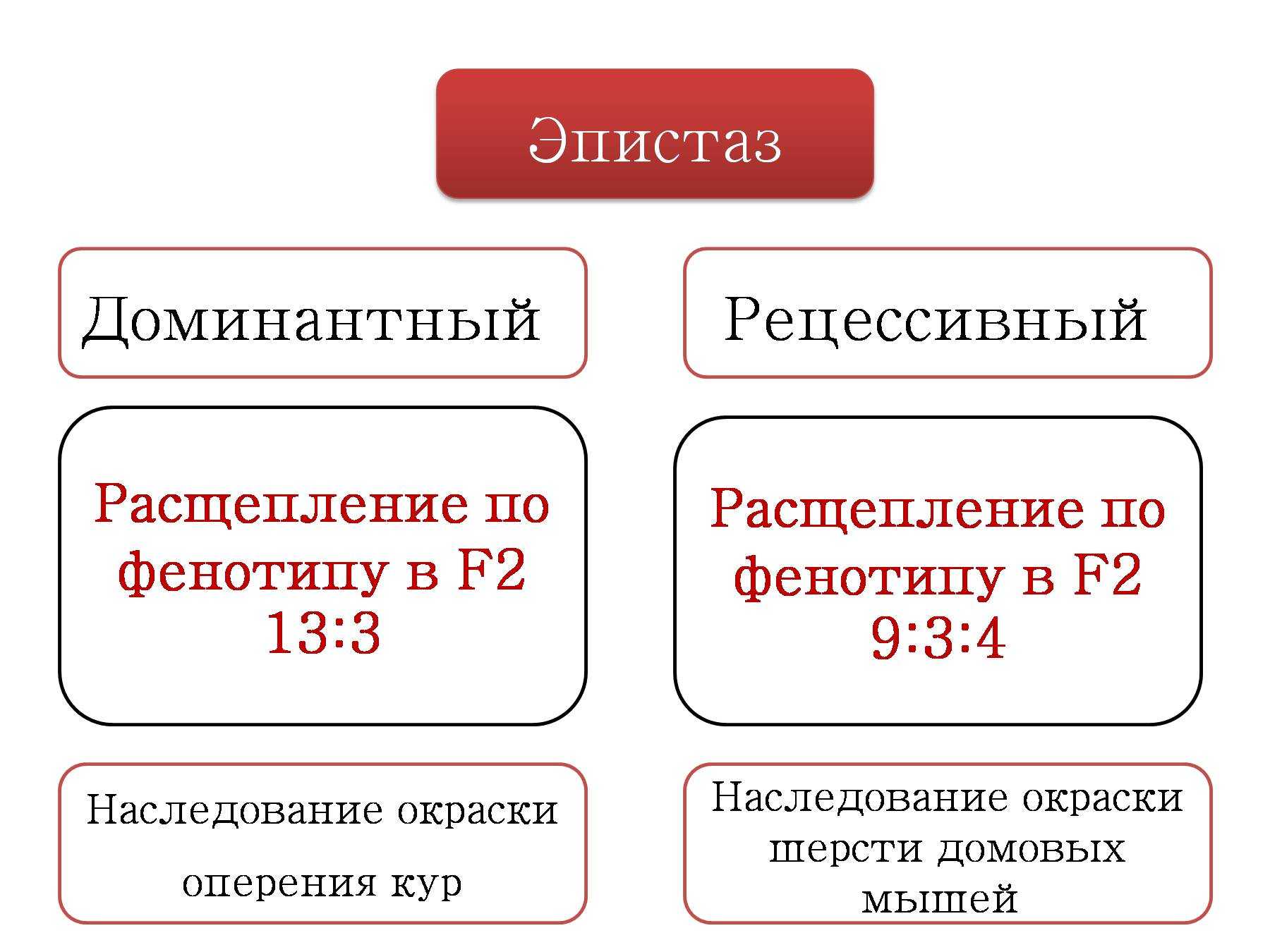
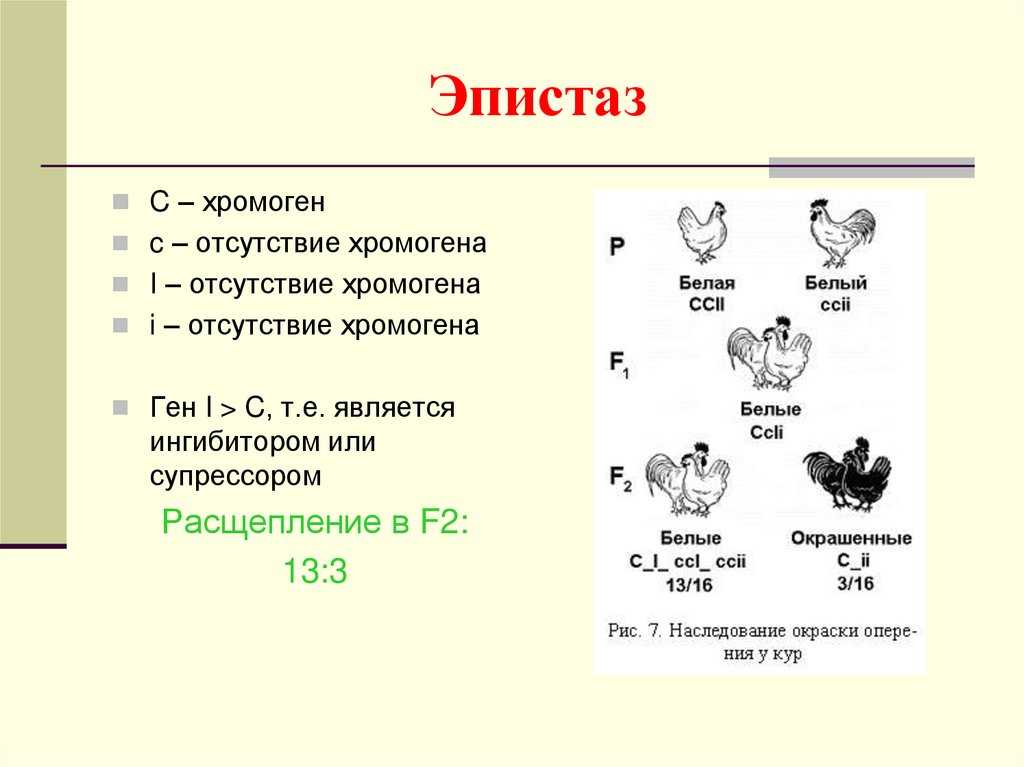
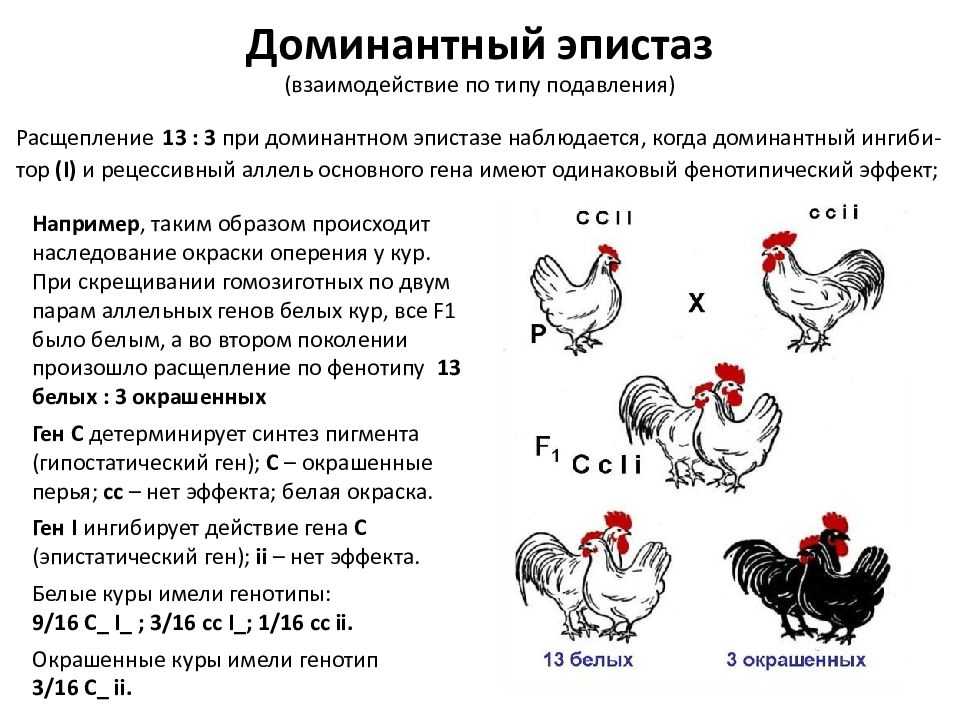
— вид взаимодействия неаллельных генов, при котором один из генов полностью подавляет действие другого гена. Ген, подавляющий действие другого гена, может называться ген-супрессор, ингибитор, эпистатичный ген. Подавляемый ген называется гипостатичным. Эпистаз может быть как доминантным, так и рецессивным. Рассмотрим пример доминантного эпистаза. У тыквы доминантный ген Y вызывает появление желтой окраски плодов, а его рецессив у — зеленой.
Кроме того, имеется доминантный ген W, подавляющий проявление любой окраски, в то время как его рецессив w не мешает окраске проявляться. Поэтому растения, имеющие в своем генотипе хотя бы один доминантный ген W, будут образовывать белые плоды независимо от того, какие аллели гена Y будут присутствовать в их генотипах.
Например, у растений с генотипами YYWW, YyWw, yyWw плоды будут белые. Эпистатическое действие рецессивного гена иллюстрирует наследование окраски шерсти у домовых мышей.
Классическое фенотипическое соотношение 9: 3: 3: 1
Эта пропорция возникает из-за комбинации анализа наследования двух разных символов. То есть это продукт сочетания двух независимых фенотипических сегрегаций (3: 1) X (3: 1).
Когда Мендель, например, анализировал высоту растения или цвет семени, каждый символ был разделен от 3 до 1. При анализе в целом, даже если они были двумя разными символами, каждый из них был отделен от 3 до 1. То есть они были распределены независимо.
Однако, когда Мендель проанализировал символы в парах, они привели к известным фенотипическим классам 9, 3, 3 и 1. Но эти классы были суммами из двух символов другой. И никогда, ни один персонаж не влиял на то, как проявился другой.
Полимерия
Это вид взаимодействия двух и более пар неаллельных генов, доминантные аллели которых однозначно влияют на развитие одного и того же признака. Полимерное действие генов может быть кумулятивным и некумулятивным. При кумулятивной полимерии интенсивность значения признака зависит от суммирующего действия генов: чем больше доминантных аллелей, тем больше степень выраженности признака. При некумулятивной полимерии количество доминантных аллелей на степень выраженности признака не влияет, и признак проявляется при наличии хотя бы одного из доминантных аллелей. Полимерные гены обозначаются одной буквой, аллели одного локуса имеют одинаковый цифровой индекс, например А1а1А2а2А3а3.
Кумулятивная полимерия имеет место при наследовании окраски зерновок пшеницы, чешуек семян овса, роста и цвета кожи человека и т.д.
| P | A1A1A2A2черные чешуйки | × | a1a1a2a2белые чешуйки |
| Типы гамет | A1A2 | a1a2 | |
| F1 | A1a1A2a2серые чешуйки, 100% | ||
| P | A1a1A2a2серые чешуйки | × | A1a1A2a2серые чешуйки |
| Типы гамет | A1A2 A1a2 a1A2 a1a2 | A1A2 A1a2 a1A2 a1a2 |
| A1A2 | A1a2 | a1A2 | a1a2 | ||
| A1A2 | A1A1A2A2черные | A1A1A1a2темно-серые | A1a1A2A2темно-серые | A1a1A2a2серые | |
| A1a2 | A1A1A2a2темно-серые | A1A1a2a2серые | A1a1A2a2серые | A1a1а2a2светло-серые | |
| a1A2 | A1a1A2A2темно-серые | A1a1A2a2серые | a1a1A2A2серые | a1a1A2a2светло-серые | |
| a1a2 | A1a1A2a2серые | A1a1a2a2светло-серые | a1a1A2a2светло-серые | a1a1a2a2желтые |
Черные чешуйки семян у овса — 1/16, темно-серые чешуйки семян у овса — 4/16, серые чешуйки семян у овса — 6/16, светло-серые чешуйки семян у овса — 4/16, желтые чешуйки семян у овса — 1/16. Расщепление по фенотипу 1:4:6:4:1.







Некумулятивная полимерия имеет место при наследовании формы плодов пастушьей сумки.
| P | A1A1A2A2треугольные | × | a1a1a2a2овальные |
| Типы гамет | A1A2 | a1a2 | |
| F1 | A1a1A2a2треугольные, 100% | ||
| P | A1a1A2a2треугольные | × | A1a1A2a2треугольные |
| Типы гамет | A1A2 A1a2 a1A2 a1a2 | A1A2 A1a2 a1A2 a1a2 |
| A1A2 | A1a2 | a1A2 | a1a2 | ||
| A1A2 | A1A1A2A2треугольные | A1A1A1a2треугольные | A1a1A2A2треугольные | A1a1A2a2треугольные | |
| A1a2 | A1A1A2a2треугольные | A1A1a2a2треугольные | A1a1A2a2треугольные | A1a1а2a2треугольные | |
| a1A2 | A1a1A2A2треугольные | A1a1A2a2треугольные | a1a1A2A2треугольные | a1a1A2a2треугольные | |
| a1a2 | A1a1A2a2треугольные | A1a1a2a2треугольные | a1a1A2a2треугольные | a1a1a2a2овальные |
Треугольная форма плодов у пастушьей сумки — 15/16, овальная форма плодов у пастушьей сумки — 1/16.
Плейотропия — множественное действие генов. Плейотропное действие генов имеет биохимическую природу: один белок-фермент, образующийся под контролем одного гена, определяет не только развитие данного признака, но и воздействует на вторичные реакции биосинтеза других признаков и свойств, вызывая их изменение.
Плейотропное действие генов впервые было обнаружено Г. Менделем, который установил, что у растений с пурпурными цветками всегда имелись красные пятна в пазухах листьев, а семенная кожура была серого или бурого цвета. То есть развитие этих признаков определяется действием одного наследственного фактора (гена).
У человека встречается рецессивная наследственная болезнь — серповидно-клеточная анемия. Первичным дефектом этой болезни является замена одной из аминокислот в молекуле гемоглобина, что приводит к изменению формы эритроцитов. Одновременно с этим возникают нарушения в сердечно-сосудистой, нервной, пищеварительной, выделительной системах. Это приводит к тому, что гомозиготный по этому заболеванию ребенок погибает в детстве. Причиной синдрома Марфана является доминантная мутация гена, контролирующего одновременно рост, длину пальцев, формирование интеллекта и форму хрусталика. Для человека с этим синдромом характерен комплекс следующих признаков — высокий рост, очень длинные гибкие («паучьи») пальцы, повышенный интеллект, близорукость.
Плейотропия широко распространена. Изучение действия генов показало, что плейотропным эффектом, очевидно, обладают многие, если не все, гены.
Таким образом, выражение «ген определяет развитие признака» в значительной степени условно, так как действие гена зависит от других генов — от генотипической среды. На проявление действия генов влияют и условия окружающей внешней среды. Следовательно, генотип является системой взаимодействующих генов.
-
Перейти к лекции №19 «Генетика пола»
-
Перейти к лекции №21 «Изменчивость»
-
Смотреть оглавление (лекции №1-25)
Эпистаз. Примеры
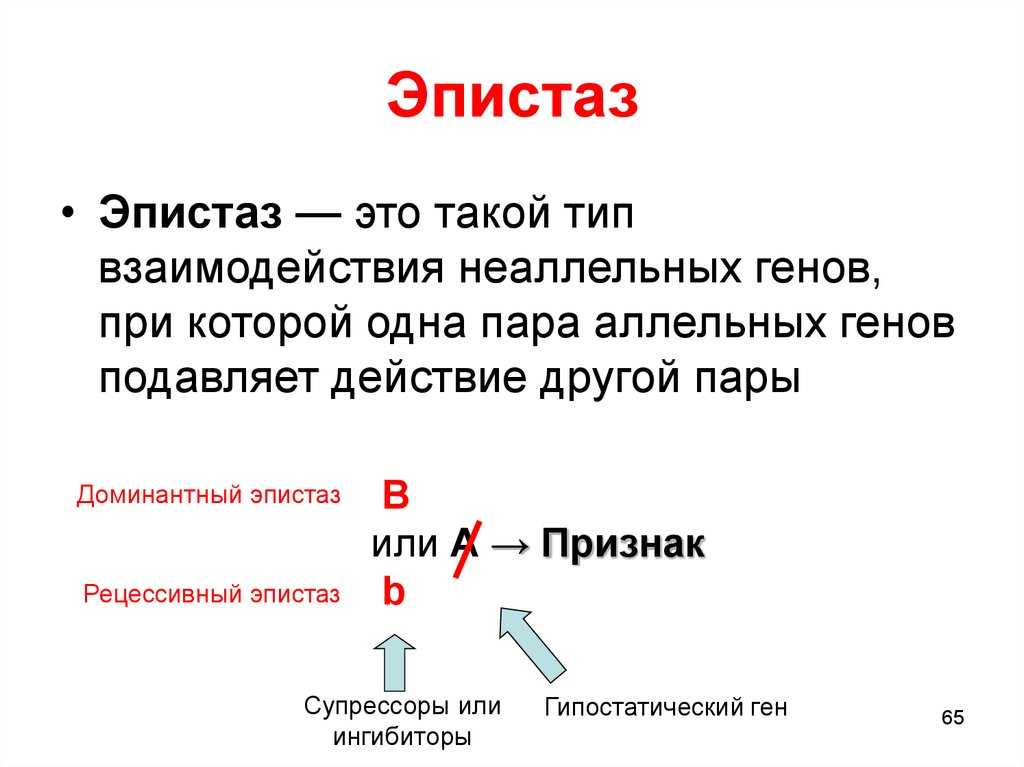
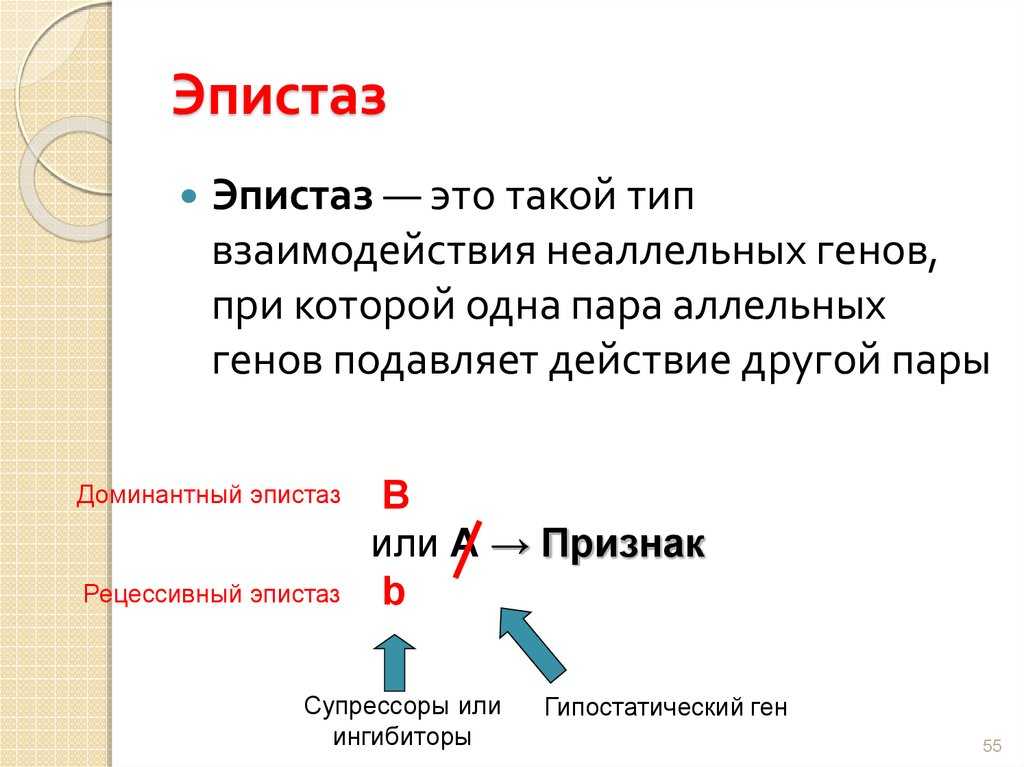
Ген называют эпистатическим (от греч. еpi — над), если его присутствие подавляет эффект какого-либо гена, находящегося в другом локусе. Эпистатические гены иногда называют ингибирующими генами, а те гены, действие которых ими подавляется, — гипостатическими.
Гены, подавляющие действие других генов, называются супрессорами или ингибиторами.
Они могут быть как доминантными, так и рецессивными. Гены-супрессоры известны у животных, растений и микроорганизмов. Обычно они обозначаются I или S.
Эпистаз принято делить на два типа: доминантный и рецессивный. Под доминантным эпистазом понимают подавление одним доминантным геном действия другого гена.
Окраска шерсти у мышей контролируется парой генов, находящихся в разных локусах.
Эпистатический ген определяет наличие окраски и имеет два аллеля: доминантный, определяющий окрашенную шерсть, и рецессивный, обусловливающий альбинизм (белая окраска). Гипостатический ген определяет характер окраски и имеет два аллеля: агути (доминантный, определяющий серую окраску) и черный (рецессивный). Мыши могут иметь серую или черную окраску в зависимости от своих генотипов, но наличие окраски возможно только в том случае, если у них одновременно имеется аллель окрашенной шерсти.
Мыши, гомозиготные по рецессивному аллелю альбинизма, будут альбиносами даже при наличии у них аллелей агути и черной шерсти. Возможны три разных фенотипа: агути, черная шерсть и альбинизм. При скрещивании можно получить эти фенотипы в различных соотношениях в зависимости от генотипов скрещиваемых особей.
Примеры эпистаза
Так, рецессивные гены окраски собак из локуса С не позволяют синтезироваться пигментам, определяющим окраску шерсти.
Собака гомозиготная по ним — белая.
Такой тип эпистаза называется рецессивным. Скрещивание белой черноносой собаки с коричневой приведет к рождению в первом поколении гетерозиготных черных собак, а скрещивание гибридов F, между собой — к рождению 9 черных, 3 коричневых и 4 белых, т. е. будет наблюдаться отклонение от нормального расщепления 9:3:3:1.
Скрещивание белой кошки, имеющей доминантный белый окрас, обусловленный доминантным аллелем W, с черной, напротив, приведет к рождению в первом поколении белых гетерозиготных котят.
Расщепление в F2 даст вместо ожидаемых 9:3:3:1 соотношение 12:3:1, где 12 — белых, 3 — полосатых и 1 черный котенок. Такой тип эпистаза называется доминантным. Отсутствие необходимых данных о роли первичных продуктов многих генов (ферментов, пептидов) в формировании сложных признаков, часто не позволяет точно установить характер взаимодействия неаллельных локусов, участвующих в биохимических процессах и составляющих основу образования этих признаков.







В одних случаях развитие признака при наличии двух неаллельных генов в доминантном состоянии рассматривают как комплементарное воздействие, в других — неразвитие признака, определяемого одним из генов при отсутствии другого гена в доминантом состоянии, расценивают как рецессивный эпистаз; если же признак развивается при отсутствии доминантного аллеля неаллельного гена, а в его присутствии не развивается, говорят о доминантом эпистазе.
Механизмы эпистаза генов
Гены – супрессоры обладают способностью подавлять действие других генов, гены ингибиторы оказывают аналогичное действие. Реакция, возникающая от такого взаимодействия, может проявиться в рецессивном и доминантном вариантах.
Доминантный эпистаз наблюдается в том случае, если доминантная аллель гена – супрессора подавляет действие другого гена. Такая аллель, как правило, обозначается буквой I.
В свою очередь рецессивный эпистаз наблюдается в диаметрально противоположном случае при угнетении одного гена другим.
Фенотипическое расщепление при каждом виде эпистаза существенно различается. При доминантном эпистазе в случае скрещивания дигетерозигот расщепление может быть в следующих соотношениях:
- тринадцать к трем и семь к шести к трем;
- двенадцать к трем и к одному.
При рецессивном эпистазе расщепление может быть в следующих соотношениях:
- девять к трем и к четырём;
- тринадцать к трем.
Взаимодействие по типу эпистаза противоположно комплементарному. При наличии комплиментарного взаимодействия наблюдается дополнение одного гена другим. Эпистаз очень схож с доминированием, но разница проявляется в участие аллеля, принадлежащего к аллеморфной паре.
При эпистазе аллель одного гена подавляет проявление аллеля из другой аллеломорфной пары, т. е. неаллельного гена. Фенотипически эпистаз выражается в отклонении от расщепления, которое ожидается от стандартного наследования. Следует отметить тот факт, что нарушения законов Менделя в данном случае не происходит, поскольку распределения аллелей всех взаимодействующих генов соответствует закону независимого комбинирования тех или иных признаков.
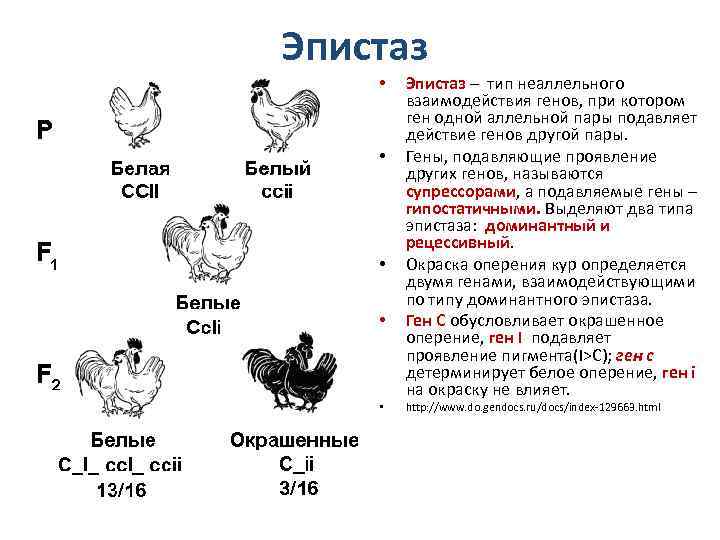
Эпистатическая система наследования обнаружена у кур. Некоторые породы кур имеют белое оперение (плимутрок, леггорн), другие породы имеют окрашенное оперение (полосатый плимутрок). При этом белое оперение различных пород кур определяется несколькими различными генами. Например, белая доминантная окраска определяется генами CCII, а рецессивная белая – ccii. Возможно и другое условное обозначение.
При этом ген С позволяет определить наличие предшественника пигмента или исследовать окрашенность пера и гены, отвечающие за не окрашенность пера. При скрещивании между собой гибридов F1 во втором поколении имеет место расщепление по окраске в отношении 13 белых: 3 окрашенных.
Также можно привести и другие примеры эпистатического взаимодействия генов. У человека существует так называемый бомбейский феномен. Люди, которые гетерозиготные по гену FUTI обладают нарушенной экспрессией генов, которые кодируют ферменты, отвечающие за образование эритроцитов A и B. На эритроцитах вместо нормальных генов образуются дефектные гены. Эти люди имеют группу крови O независимо от аллелей генов, которые определяют тип группы крови по системе ABO, не имеют антиегна A, антигена B.
Гены, которые определяют окраску шерсти у мышей также обладают эпистатическим взаимодействием. Окраска агаути у мышей обусловлена чередованием кольцевых пигментных полос по длине каждой части волосяного покрова. Она доминирует над чёрной окраской и определяется рецессивным мутантным аллелем. Если рецессивная мутация имеется в другом локусе, то пигментация полностью исчезает, и эта ситуация совершенно не зависит от того, какими аллелями представлен ген A.
Доминантный эпистаз у тыквы проявляется в окраске плодов. Доминантный аллель тыквы обуславливает ее белую окраску, вне зависимости от того, какими аллелями представлен ген B. Если локус A представлен рецессивной аллелью, то растения с генотипами BB и Bb имеют жёлтые плоды, а bb — зелёные.
Встречаются ситуации, когда наблюдается двойной эпистаз. Он встречается в тех случаях, когда встречаются гены, кодирующие ферменты одного метаболического пути. Если внутри клетки не будет создаваться нормальная форма одного из ферментов, то также не будет образовываться промежуточный продукт, который является фундаментом для всех последующих стадий. В результате нормативного продукта не будет образовываться при любой комбинации аллелей.
Также в качестве примера можно привести гены мухи – дрозофилы, которые кодируют ферменты – катализаторы, участвующие в превращении триптофана, который приобретает бурую окраску во время химических превращений. Если в процессе будет участвовать только рецессивный ген, то фенотип мух по окраске будет существенно отличаться от дикого типа.
Таким образом, эпистатическое взаимодействие неаллельных генов является одним из примеров подавления одного гена другим. Он позволяет отследить закономерности наследования генов и отразить специфику их взаимодействия внутри генотипа.
На модификацию дигибридного скрещивания при рецессивном эпистазе
В брак вступают супруги, имеющие кровь II группы, матери которых имели I группу крови, и гетерозиготные по гену рецессивного эпистаза. Определите вероятные группы крови детей с учетом рецессивного эпистаза?
Решение
Обозначим гены:
| Ген | Признак |
| I0- 1А- 1В- 1А1В- хх>1А- хх>1в- хх>1А1в- X — х — | Iгруппа крови II группа крови III группа крови IV группа крови I гр. кр. I гр. кр. Рецессивный эпистаз I гр. кр. нормальный ген рецессивный эпистатический ген. |
Определите вероятные группы крови детей с учетом рецессивного эпистаза?
Фенотипы родителей:
II гр. крови — II гр. крови
гетерозигот. по 2 генам гетерозигот. по 2 генам







Ответ:_Расщепление по фенотипу 9:7, вероятность рождения со второй группой крови составляет 9/16, с первой 7/16, в 3/16 (IAIAcx, IAI°xx, IAI°xx) проявилась I группа крови вследствие рецессивного эпистаза и в 4/16 за счет сочетания гамет в зиготе (см. решетку Пеннета).
Явление эпистаза генов
Необходимо ответить тот факт, что очень редко развитие того или иного признака определяется одним геном. В большинстве случаев какой – либо ген в единственном экземпляре становится фундаментом для развития различных признаков. Такое явление называется плейотропным взаимодействием генов. Также весьма распространено такое явление, когда один признак контролируется несколькими генами.
Определение 2
Неаллельные гены – это гены, которые расположены в различных местах (участках хромосом) и кодируют разные белки.
Неаллельные гены могут взаимодействовать между собой, когда один признак проявляется под действием нескольких генов.
В науке выделяется три формы взаимодействия генов между собой:
- комплементарность (или такое взаимодействие генов, которые дополняют друг друга и обуславливают развитие нового признака);
- полимерия или процесс генетического взаимодействия, при котором неаллельные гены влияют в одном напралвении на развитие одинакового признака;
- эпистаз (подавление гена иным геном).
Все указанные типы неаллельных взаимодействий встречаются с равной степенью вероятности и должны оцениваться с позиций гибридологического подхода.
Методы и модельные системы
Регрессионный анализ
Количественная генетика фокусируется на генетической изменчивости из-за генетических взаимодействий. Любые два локусных взаимодействия с определенной частотой гена можно разложить на восемь независимых генетических эффектов с помощью взвешенной регрессии . В этой регрессии наблюдаемые генетические эффекты двух локусов рассматриваются как зависимые переменные, а «чистые» генетические эффекты используются как независимые переменные. Поскольку регрессия является взвешенной, распределение между компонентами дисперсии будет изменяться в зависимости от частоты генов. По аналогии можно расширить эту систему до трех или более локусов или до цитоядерных взаимодействий.
Двойные мутантные циклы
При анализе эпистаза в гене можно использовать сайт-направленный мутагенез для генерации различных генов, а их белковые продукты могут быть проанализированы (например, на стабильность или каталитическую активность). Иногда это называют двойным мутантным циклом, и он включает производство и анализ белка дикого типа, двух одиночных мутантов и двойного мутанта. Эпистаз измеряется как разница между совокупными эффектами мутаций и суммой их индивидуальных эффектов. Это можно выразить как свободную энергию взаимодействия. Ту же методологию можно использовать для исследования взаимодействий между более крупными наборами мутаций, но все комбинации должны быть получены и проанализированы. Например, существует 120 различных комбинаций из 5 мутаций, некоторые или все из которых могут проявлять эпистаз …
Вычислительный прогноз
Были разработаны многочисленные вычислительные методы для обнаружения и характеристики эпистаза. Многие из них полагаются на машинное обучение для обнаружения неаддитивных эффектов, которые могут быть упущены статистическими подходами, такими как линейная регрессия. Например, многофакторное снижение размерности (MDR) было разработано специально для непараметрического и безмодельного обнаружения комбинаций генетических вариантов, которые предсказывают фенотип, такой как статус заболевания, в популяциях людей . . Некоторые из этих подходов широко рассмотрены в литературе. Еще совсем недавно было показано , что методы, использующие идеи теоретической информатики ( преобразование Адамара ) или вывод максимального правдоподобия, позволяют отличать эпистатические эффекты от общей нелинейности в структуре карты генотип-фенотип, в то время как другие использовали анализ выживаемости пациентов для выявления нестандартных факторов. линейность.
Полное доминирование
Полное доминирование — это вид взаимодействия аллельных генов, при котором фенотип гетерозигот не отличается от фенотипа гомозигот по доминанте, то есть в фенотипе гетерозигот присутствует продукт доминантного гена. Полное доминирование широко распространено в природе, имеет место при наследовании, например, окраски и формы семян гороха, цвета глаз и цвета волос у человека, резус-антигена и мн. др.
Наличие резус-антигена (резус-фактора) эритроцитов обусловливается доминантным геном Rh. То есть генотип резус-положительного человека может быть двух видов: или RhRh, или Rhrh; генотип резус-отрицательного человека — rhrh. Если, например, мать — резус-отрицательная, а отец резус-положительный и гетерозиготен по этому признаку, то при данном типе брака с одинаковой вероятностью может родиться как резус-положительный, так и резус-отрицательный ребенок.
| Р | rhrhрезус-отрицательная | × | Rhrhрезус-положительный |
| Типы гамет | rh | Rh rh | |
| F | Rhrhрезус-положительный50% | rhrhрезус-отрицательный50% |
Между резус-положительным плодом и резус-отрицательной матерью может возникнуть резус-конфликт.
ссылки
- Брукер Р.Дж. (2017). Генетика: анализ и принципы. McGraw-Hill Higher Education, Нью-Йорк, Нью-Йорк, США.
- Goodenough, U. W. (1984) Genetics. W. B. Saunders Co. Ltd, Пкиладелия, Пенсильвания, США.
- Griffiths A.J.F., Wessler R., Carroll S.B., Doebley J. (2015). Введение в генетический анализ (11-е изд.). Нью-Йорк: У. Х. Фриман, Нью-Йорк, Нью-Йорк, США.
- Miko, I. (2008) Epistasis: взаимодействие генов и фенотипические эффекты. Природоведческое образование 1: 197. nature.com
- Уайт Д., Рабаго-Смит М. (2011). Генотип-фенотипические ассоциации и цвет глаз человека. Журнал генетики человека, 56: 5-7.
- .