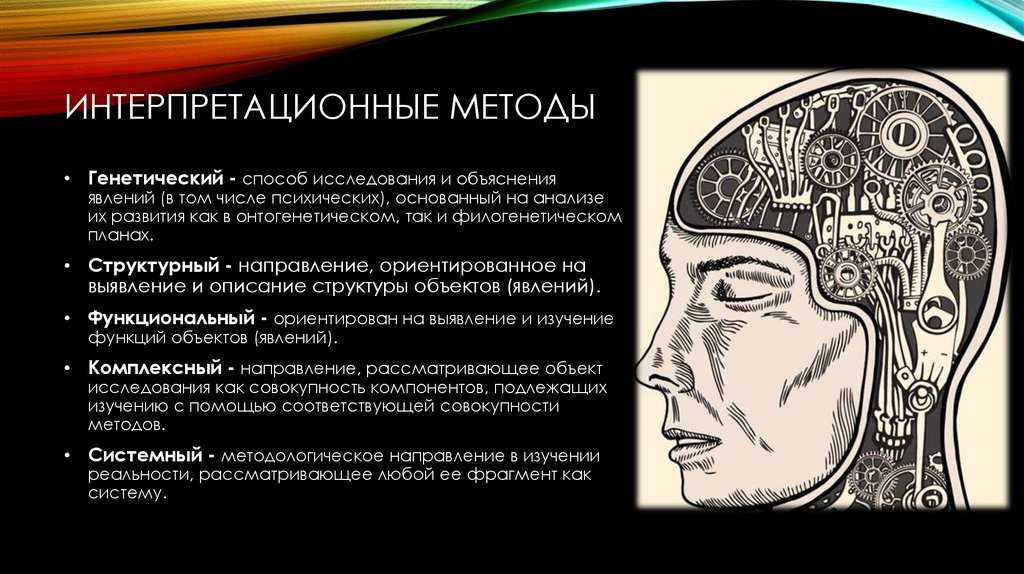
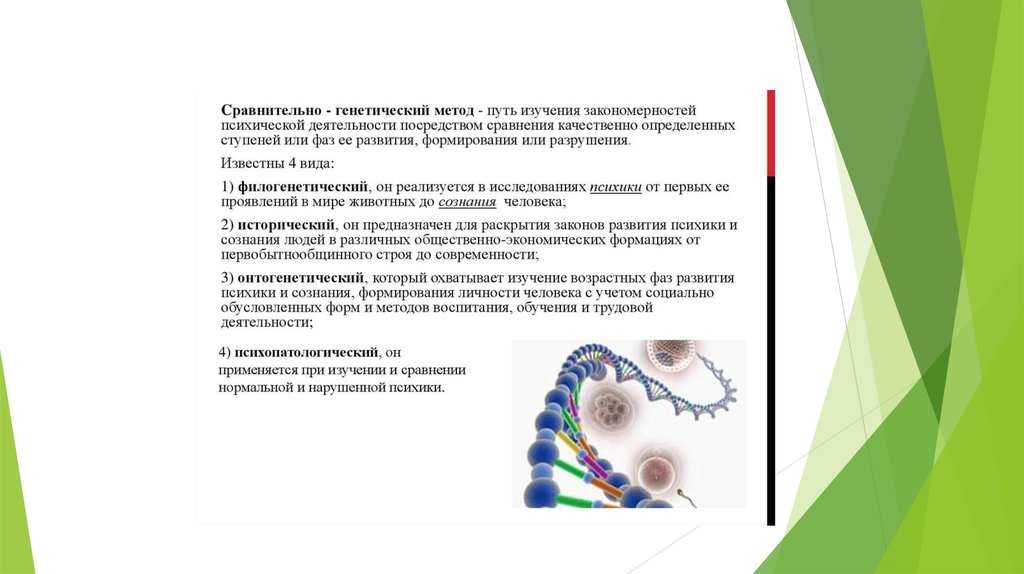
Критика филогенетической классификации
С кладистской точки зрения эволюционная систематика субъективно полагается на наиболее видимые сходства между видами как основу для классификации, которая не отражает правильно родственные отношения между видами. Эта критика отвергается эволюционными систематиками, которые настаивают на заинтересованности парафилетических групп в правильном представлении эволюционных процессов. Некоторые исследователи, эволюционные систематики , критикуют результат филогенетической классификации, потому что клады имеют сильную тенденцию группировать виды, которые не похожи друг на друга, и, наоборот, разделять виды, которые похожи друг на друга. В основном это связано с очень большой неоднородностью скорости эволюции от одной ветви к другой. Поэтому ставятся под сомнение не методы построения клад, а их использование для создания формальной классификации. Согласно этой школе, классификация должна отражать не только топологию древа жизни, но и эволюционные дистанции, что равносильно разрешению парафилетических групп .
Анализ генома для составления филогенетических рядов
Для анализа генома используются различные методы, включая секвенирование ДНК и биоинформатический анализ. Секвенирование ДНК позволяет определить последовательность нуклеотидов в геноме организма, включая гены, регуляторные регионы и другие необходимые элементы. Эта информация затем используется для сравнения геномов различных организмов и определения их филогенетических родственных связей.
Биоинформатический анализ генома включает в себя обработку и анализ больших объемов данных, полученных в результате секвенирования. В процессе анализа используются различные программы и базы данных, позволяющие сравнивать и аннотировать гены, определять эволюционно-консервативные участки генома и проводить другие вычислительные операции. Результаты анализа генома могут быть представлены в виде филогенетического дерева, показывающего относительные родственные связи между организмами и их эволюционное развитие.
Анализ генома является мощным инструментом для изучения эволюции организмов и определения их филогенетического положения. Этот подход позволяет расширить наше понимание о разнообразии жизни на Земле и выявить новые филогенетические связи, которые могут быть скрыты на основе только морфологических или анатомических признаков.
Филогенетический обобщенный метод наименьших квадратов (PGLS)
Вероятно, наиболее часто используемым PCM является филогенетический обобщенный метод наименьших квадратов (PGLS). Этот подход используется для проверки наличия взаимосвязи между двумя (или более) переменными с учетом того факта, что происхождение не является независимым. Метод является частным случаем обобщенный метод наименьших квадратов (GLS) и, как таковая, оценка PGLS также беспристрастный, последовательный, эффективный, и асимптотически нормальный. Во многих статистических ситуациях, когда GLS (или, обыкновенный метод наименьших квадратов ) используется остаточная ошибка ε считаются независимыми и одинаково распределенными случайными величинами, которые считаются нормальными
- ε∣Икс∼N(,σ2яп).{displaystyle varepsilon mid Xsim {mathcal {N}} (0, sigma ^ {2} I_ {n}).}
тогда как в PGLS предполагается, что ошибки распределяются как
- ε∣Икс∼N(,V).{displaystyle varepsilon mid Xsim {mathcal {N}} (0, mathbf {V}).}
куда V представляет собой матрицу ожидаемой дисперсии и ковариации остатков с учетом эволюционной модели и филогенетического дерева. Следовательно, это структура остатков, а не сами переменные, которые показывают филогенетический сигнал. Это уже давно является источником путаницы в научной литературе. Предложен ряд моделей структуры V Такие как Броуновское движениеОрнштейн-Уленбек, и λ-модель Пагеля. (Когда используется модель броуновского движения, PGLS идентичен независимой оценке контрастов.). В PGLS параметры эволюционной модели обычно оцениваются совместно с параметрами регрессии.
PGLS может применяться только к вопросам, в которых зависимая переменная непрерывно распространяется; однако филогенетическое древо также может быть включено в остаточное распределение обобщенные линейные модели, что позволяет обобщить подход на более широкий набор распределений для ответа.
Методы составления филогенетических рядов: сравнение достоинств и недостатков
Один из основных вопросов в биологии – исследование взаимосвязей между организмами и их эволюционные процессы. Для составления филогенетического ряда организмов используются различные методы, каждый из которых имеет свои достоинства и недостатки. В данной статье мы рассмотрим несколько из них.
-
Метод фенетической классификации
Данный метод основан на сравнении фенотипических признаков организмов. Фенотипический признак — это видимое проявление генетической информации в организме. Однако, этот метод не учитывает эволюционную историю и не позволяет определить филогенетические отношения между организмами.
-
Молекулярно-генетический метод
Данный метод основан на сравнении генетической информации организмов, такой как последовательность ДНК или РНК. Он позволяет установить более точные филогенетические отношения между организмами. Однако, этот метод требует наличия большого количества генетических данных и их анализа, что может быть затруднительно и затратно.
-
Морфологический метод
Данный метод основан на сравнении морфологических признаков организмов, таких как форма, цвет, размер и структура органов. Он является относительно простым и доступным для применения, однако может не учитывать некоторые эволюционно-значимые признаки и может оказаться подвержен влиянию окружающей среды.
В заключение, каждый из приведенных методов имеет свои преимущества и ограничения. Их выбор зависит от конкретной задачи и доступности данных. Часто, для более точного определения филогенетических рядов, рекомендуется использовать комбинацию различных методов.
Филогенетический обобщенный метод наименьших квадратов (PGLS)
Вероятно, наиболее часто используемым PCM является филогенетический обобщенный метод наименьших квадратов (PGLS). Этот подход используется для проверки наличия взаимосвязи между двумя (или более) переменными с учетом того факта, что происхождение не является независимым. Этот метод является частным случаем обобщенных наименьших квадратов (GLS), и поэтому оценка PGLS также несмещена , согласована , эффективна и асимптотически нормальна . Во многих статистических ситуациях, когда используется GLS (или обычный метод наименьших квадратов ), остаточные ошибки ε считаются независимыми и одинаково распределенными случайными величинами, которые считаются нормальными.
- ε∣Икс∼N(,σ2яп).{\ displaystyle \ varepsilon \ mid X \ sim {\ mathcal {N}} (0, \ sigma ^ {2} I_ {n}).}
тогда как в PGLS предполагается, что ошибки распределяются как
- ε∣Икс∼N(,V).{\ displaystyle \ varepsilon \ mid X \ sim {\ mathcal {N}} (0, \ mathbf {V}).}
где V — матрица ожидаемой дисперсии и ковариации остатков с учетом эволюционной модели и филогенетического дерева. Следовательно, филогенетический сигнал показывает структура остатков, а не сами переменные . Это уже давно является источником путаницы в научной литературе. Для структуры V был предложен ряд моделей, таких как броуновское движение Орнштейна-Уленбека и λ-модель Пагеля. (Когда используется модель броуновского движения, PGLS идентичен независимой оценке контрастов.) В PGLS параметры эволюционной модели обычно оцениваются совместно с параметрами регрессии.
PGLS может применяться только к вопросам, в которых зависимая переменная непрерывно распределяется; однако филогенетическое дерево также может быть включено в остаточное распределение обобщенных линейных моделей , что позволяет обобщить подход к более широкому набору распределений для ответа.
Палеонтологические данные и филогенетические ряды
Палеонтологические данные играют важную роль в составлении филогенетических рядов организмов. Они позволяют ученые реконструировать историю эволюции различных видов и создавать систематические классификации.
Основным источником палеонтологических данных являются ископаемые останки, которые могут быть найдены в разных геологических отложениях. Ископаемые останки организмов представляют собой кости, зубы, скелеты или другие останки, которые сохранились до наших дней. По ископаемым останкам ученые могут определить внешний вид, морфологию, анатомию и другие характеристики древних организмов.
На основе палеонтологических данных строятся филогенетические ряды, которые показывают эволюционные отношения между различными видами. Филогенетические ряды отображают общую историю жизни на Земле и позволяют ученым понять, как разные виды связаны между собой в эволюционном смысле.
Филогенетические ряды строятся на основе сравнительного анализа анатомических, морфологических и генетических данных. Палеонтологические данные дополняют этот анализ, предоставляя ученым информацию о древних организмах, которые не сохранились в современном мире. Это позволяет ученым восстановить более полное представление об эволюции организмов и уточнить их филогенетические ряды.









Для строительства филогенетических рядов, палеонтологические данные анализируются в сочетании с данными из других областей науки, таких как молекулярная биология и морфология. Такой мультидисциплинарный подход позволяет ученым получить наиболее точное представление о происхождении и эволюции разных видов на протяжении миллионов лет.
Использование палеонтологических данных в составлении филогенетических рядов позволяет ученым проследить динамику эволюции истории жизни на Земле и понять, какие процессы привели к появлению различных видов. Это важный шаг в изучении биологического разнообразия и понимании пути, по которому оно развивалось на протяжении миллионов лет.
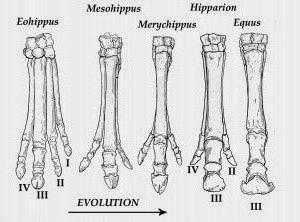
Промежуточные звенья
Порядка 25 миллионов лет назад климат на планете изменился, и на смену лесам стали приходить степные просторы. В миоцене (20 миллионов лет назад) появляются мезогиппус и парагиппус, уже более похожие на современных лошадей. А первым травоядным предком в филогенетическом ряду лошади принято считать мерикгиппуса и плиогиппуса, которые выходят на арену жизни 2 миллиона лет назад. Гиппарион — последнее трехпалое звено
Этот предок жил в миоцене и плиоцене на равнинах Северной Америки, Азии и Африки. Эта трехпалая лошадка, напоминающая газель, еще не имела копыт, но могла быстро бегать, ела траву и именно она заняла огромные территории.
Типы филогенетических деревьев[]
Рис. 1: Теоретически укоренённое дерево для генов рРНК
Рис. 2: Неукоренённое дерево для семейства супергена миозина
Укоренённое дерево — дерево, содержащее выделенную вершину — корень. Корневое дерево можно считать ориентированным графом, поскольку на нем имеется естественная ориентация — от корня к листьям. Каждый узел корневого дерева отвечает последнему общему предку нижележащих листьев дерева. Рисунок 1 представляет корневое филогенетическое дерево, окрашенное в соответствии с трёхдоменной системой живых организмов. .
Неукоренённое дерево не содержит корня и отражает связь листьев без предполагаемого положения общего предка. Необходимость рассматривать некорневые деревья возникает из-за того, что часто связи между узлами восстановить легче, чем направление эволюции. Рисунок 2 иллюстрирует некорневое филогетическое дерево. . Наиболее достоверным методом для превращения неукорененного дерева в укорененное (для этого надо либо объявить корнем один из узлов, либо разбить одну из ветвей на две, выходящие из корня) является использование достоверной «внешней группы» видов — достаточно близких к интересующему нас набору видов для достоверного восстановления топологии дерева для объединенного множества видов, но в то же время заведомо являющихся отдельной группой. Иногда положение корня можно угадать, исходя из каких-либо дополнительных знаний о природе изучаемых объектов (видов, белков, etc.)
Укоренённое и неукоренённое филогетическое дерево может быть бифуркационным или небифуркационным, а также маркированным или немаркированным.
В бифуркационном дереве к каждому узлу подходят ровно три ветви (в случае корневого дерева — одна входящая ветвь и две исходящие). Таким образом бифуркационное дерево предполагает, что все эволюционные события состояли в происхождении от предкового объекта ровно двух потомков. К узлу небифуркационного дерева могут подходить четыре и более ветви.
Маркированное дерево содержит названия листьев, тогда как немаркированное просто отражает топологию.
Дендрограмма — общий термин, обозначающий схематическое представление филогенетического дерева.
Кладограмма — филогенетическое дерево, не содержащее информации о длинах ветвей.
Филограмма (или фенограмма) — филогенетическое дерево, содержащее информацию о длинах ветвей; эти длины представляют изменение некой характеристики.
Хронограмма — филограмма, длины ветвей в которой представляют эволюционное время.
Дикие лошади
Сегодня настоящих диких лошадей практически не осталось. Российский тарпан считается вымершим видом, а лошадь Пржевальского в естественных условиях не встречается. Табуны лошадей, которые пасутся вольно, — это одичавшие одомашненные формы. Такие лошади хоть и быстро возвращаются к дикой жизни, но все же отличаются от истинно диких лошадей.
У них длинные гривы и хвосты, и они разномастные. Исключительно буланые лошади Пржевальского и мышастые тарпаны имеют как бы подстриженные челки, гривы и хвосты.

В Центральной и Северной Америке дикие лошади были полностью истреблены индейцами и появились там только после прибытия европейцев в XV веке. Одичавшие потомки лошадей конкистадоров дали начало многочисленным табунам мустангов, численность которых сегодня контролируется отстрелом.
Кроме мустангов в Северной Америке есть два вида диких островных пони — на островах Ассатиг и Сейбл. Полудикие табуны лошадей камаргу встречаются на юге Франции. В горах и болотах Британии тоже можно встретить некоторых диких пони.

Принципы
Критика традиционных классификаций
Сравнение классической и филогенетической классификаций. В последнем группа рыб не существует. А «рыба» целаканта ближе к человеку, чем к форели , как показывают филогенетические исследования .
В традиционной классификации не всегда были направлены на след родстве и эволюции видов. Существовали научные классификации в доэволюционном контексте. Однако в соответствии с этой классификацией можно предположить, что рыба всегда будет ближе к другой рыбе, чем к другому, не относящемуся к рыбам виду. На самом деле это не всегда так. Действительно, для рыб характерны чешуя и плавники. Однако люди разделяют с некоторыми видами рыб, такими как целаканты , другой характер: мясистая и неизлучаемая конечность. Например, этот последний признак отсутствует у форели. Латимерия ближе к человеку или к форели? Что мы должны использовать для установления ближайшего родства, мясистый член или наличие плавников?







![Глава 2 от синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции. логика случая [о природе и происхождении биологической эволюции]](https://lesniepolyani.ru/wp-content/uploads/6/2/d/62dfc1f4f042f87e6a7f6f8a31d6823d.jpeg)
Эволюционные исследования показали, что определенные черты эволюционировали, чтобы трансформироваться. Так обстоит дело с рыбьим плавником, который превратился в ходячую конечность у четвероногих , таких как люди. Классическая классификация, использующая признак «плавник», исключает людей из группы, которая демонстрирует плавники, хотя этот признак присутствует, но в развитой форме. То же самое и с такими наследственными признаками, как наличие чешуи (которая исчезла у некоторых видов) или гидродинамическая форма тела. Классическая классификация, использующая наиболее заметные признаки, не позволяет правильно оценить степень родства между видами. Однако эта классификация полезна для распознавания видов по ключам определения или для управления биологическими коллекциями.
Организация в клады
Каждая группа или клада должны соответствовать одному и тому же определению: клада включает всех потомков предка и самого предка. Мы также говорим о монофилетической группе . Таким образом, виды одной клады всегда будут ближе друг к другу, чем к другому виду вне этой клады. Это не относится к некоторым группам классической классификации, таким как рыбы, которые не образуют кладу.
Определенные группы, такие как млекопитающие, представленные в классической классификации, действительно составляют клады; эти группы сохранились в филогенетической классификации. Представители клады демонстрируют по крайней мере одну производную характеристику, характерную для всей группы, такую как наличие вымени млекопитающих. Это не относится к рыбам, рептилиям, которые представляют наследственные признаки, и производным (эволюционировавшим) признакам, но которые не являются исключительными для них (пример: наличие пальцев для рептилий). Производный символ был унаследован от одного и того же общего предка.
Филогенетически обоснованные компьютерные симуляции Монте-Карло
Данные для признака с непрерывной оценкой могут быть смоделированы таким образом, что таксоны на концах гипотетического филогенетического дерева будут демонстрировать филогенетический сигнал, то есть близкородственные виды будут иметь тенденцию походить друг на друга.
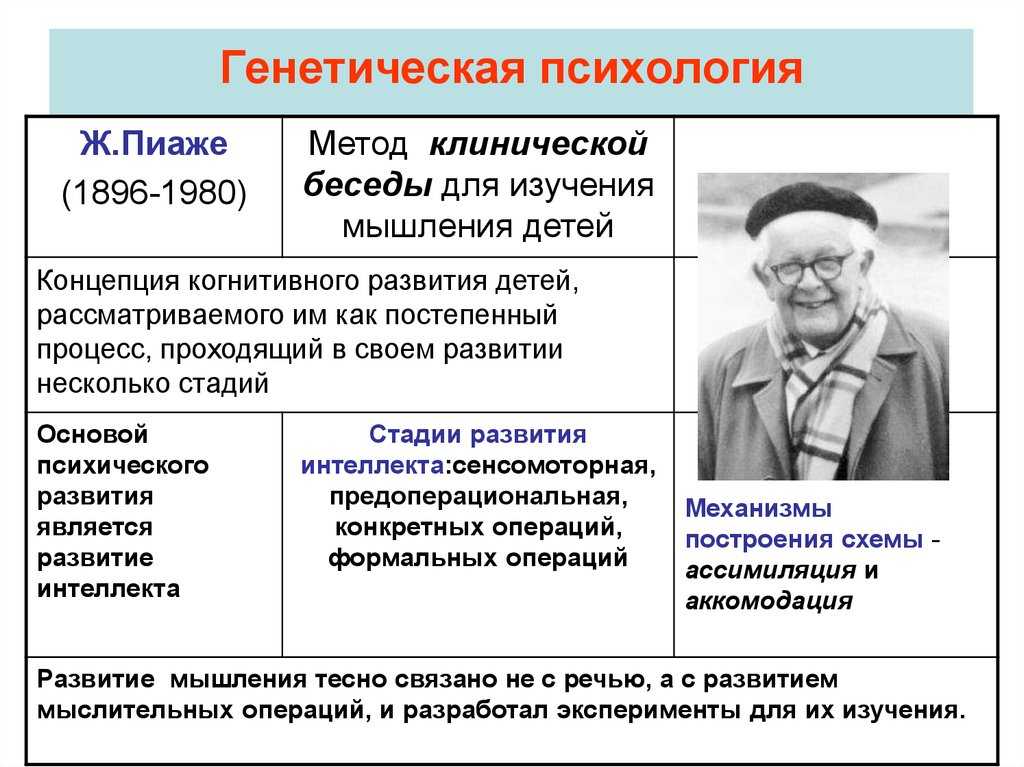
Мартинс и Гарланд предложили в 1991 году, что одним из способов учета филогенетических отношений при проведении статистического анализа было использование компьютерного моделирования для создания множества наборов данных, которые согласуются с проверяемой нулевой гипотезой (например, отсутствие корреляции между двумя признаками, отсутствие различий между двумя экологически определенными группами видов), но они имитируют эволюцию по соответствующему филогенетическому дереву. Если такие наборы данных (обычно 1000 или более) анализируются с помощью той же статистической процедуры, которая используется для анализа реального набора данных, то результаты для смоделированных наборов данных можно использовать для создания филогенетически правильных (или «ПК»).) нулевые распределения тестовой статистики (например, коэффициент корреляции, t, F). Такие подходы к моделированию также можно комбинировать с такими методами, как филогенетически независимые контрасты или PGLS (см. Выше).
Филогенетический анализ: как определить эволюционные связи
Существует несколько методов для составления филогенетических рядов организмов. Один из наиболее распространенных методов — это анализ молекулярных данных. Исследователи анализируют генетические последовательности ДНК или РНК разных организмов, чтобы определить изменения, которые произошли в их генетическом материале в результате эволюции. Более похожие последовательности генов указывают на то, что организмы более тесно связаны эволюционно.
Другой метод — это морфологический анализ. Он основывается на изучении внешнего вида и строения организмов. Исследователи анализируют анатомические, физиологические и поведенческие характеристики организмов, чтобы определить их эволюционные связи. Более схожие признаки у организмов указывают на их близкие эволюционные отношения.
Также, современные методы филогенетического анализа включают в себя комбинирование молекулярных и морфологических данных, а также использование компьютерных алгоритмов и моделей для более точного определения филогенетических связей между организмами.
В целом, филогенетический анализ является важным инструментом в изучении эволюции жизни на Земле. Он позволяет исследователям понять, как различные организмы связаны между собой и как они развивались со временем. Это позволяет лучше понять механизмы эволюции и ее последствия для различных видов и экосистем в целом.
Методы составления филогенетических рядов организмов
Филогенетика — это наука, изучающая эволюционные отношения между организмами. Филогенетические ряды организмов представляют собой группы организмов, классифицированных по их эволюционным связям
Составление филогенетических рядов является важной задачей для понимания эволюции жизни на Земле и классификации организмов
Молекулярные методы
Молекулярные методы составления филогенетических рядов основаны на сравнении генетических последовательностей организмов. Один из наиболее распространенных методов — это анализ последовательности рибосомных РНК (рРНК). Рибосомная РНК является специальным типом РНК, которая участвует в синтезе белка. Ее последовательность изменяется в течение эволюции, и сравнение последовательностей рибосомной РНК разных организмов позволяет определить их степень родства.
Другой молекулярный метод — это анализ последовательности ДНК-кода определенных генов. Код ДНК задает последовательность аминокислот в белках, и сравнение кодов разных генов позволяет определить их эволюционные отношения. Один из часто используемых генов — это ген цитохрома b, который участвует в процессе дыхания.
Морфологические методы
Морфологические методы основаны на анализе морфологических признаков организмов. Один из таких методов — это сравнение анатомических структур организмов. Например, сравнение структуры кости у разных видов позволяет установить их эволюционные отношения.
Другой морфологический метод — это сравнение формы и структуры органов организмов. Например, сравнение формы листьев у разных видов растений может указывать на их родство.
Филогенетические деревья
Результатом применения различных методов к составлению филогенетических рядов являются филогенетические деревья. Филогенетическое дерево представляет собой графическую диаграмму, на которой показаны эволюционные отношения между организмами. На дереве каждый узел представляет собой общего предка, и расстояние между узлами отражает степень родства. Ветви дерева представляют собой эволюционные линии разных организмов, а их взаимное расположение показывает временной порядок эволюционных событий.
Филогенетические деревья могут быть очень сложными и содержать сотни или тысячи организмов. Они играют важную роль в сравнительной геномике, эволюционной биологии и классификации организмов.
Приложения
Филогенетические сравнительные подходы могут дополнять другие способы изучения адаптации, такие как изучение природных популяций, экспериментальные исследования и математические модели. Межвидовые сравнения позволяют исследователям оценить общность эволюционных явлений, рассматривая независимые эволюционные события. Такой подход особенно полезен, когда внутри видов мало или совсем нет изменений. А поскольку их можно использовать для явного моделирования эволюционных процессов, происходящих в течение очень длительных периодов времени, они могут дать представление о макроэволюционных вопросах, которые когда-то были исключительной областью палеонтология.
Домашний ассортимент площадей 49 видов млекопитающие в отношении их размер тела. Виды с более крупным телом, как правило, имеют более крупный ареал обитания, но при любом заданном размере тела представители отряда Хищник (плотоядные животные и всеядные ), как правило, имеют большие домашние диапазоны, чем копытные (все из которых травоядные животные ). Считается ли эта разница статистически значимой, зависит от того, какой тип анализа применяется.
Семенники масса различных видов Приматы в отношении их размер тела и система спаривания. Виды с более крупным телом, как правило, имеют более крупные семенники, но у видов любого размера тела, у которых самки, как правило, спариваются с несколькими самцами, есть самцы с более крупными семенниками.
Филогенетические сравнительные методы обычно применяются к таким вопросам, как:
Что склон из аллометрический масштабирование отношения?
→ Пример: как масса мозга зависит от массы тела ?
Делай разные клады организмов различаются по некоторым фенотипический черта?
→ Пример: делать псовые иметь сердце больше, чем кошачьи ?
Делайте группы видов, которые разделяют поведенческий или экологическая особенность (например, социальная система, рацион питания ) отличаются средним фенотипом?
→ Пример: у хищников больше домашние диапазоны чем травоядные?
Что это было наследственный состояние черта ?
→ Пример: откуда взяли эндотермия эволюционируют в линию, которая привела к млекопитающим?
→ Пример: где, когда и почему плаценты и живородство эволюционировать?
Подает ли признак значительный филогенетический сигнал в определенной группе организмов? Некоторые типы черт склонны «следовать филогенезу» больше, чем другие?
→ Пример: поведенческие черты более лабильный во время эволюции?
Есть ли видовые различия в особенности истории жизни компромисс, как в так называемом континууме быстро-медленно?
→ Пример: почему у мелкотелых видов короче продолжительность жизни чем их более крупные родственники?