

Гнездостроение
Начнем повествование о простейшем подземном гнезде, которое
представляет собой наклонный ход с расширением,в конце (Alloformica aberrans
) или одну камеру, часто располагающуюся под камнем, под корой дерева или
под его упавшим стволом (Leptothorax). Однако у большинства видов гнезда
представляют собой сложную систему ходов (галерей) и камер. Ходы, как правило,
имеют в сечении форму круга, и их диаметр у разных видов варьирует от 1 до
10 мм. Камеры у большинства видов располагаются горизонтально и в плане обычно
имеют форму овала или круга. Пол камер, как правило, плоский и выглаженный,
потолок либо параллелен полу, либо сводчатый. У мелких муравьев часто камеры
имеют неправильную форму. В этом случае личинки не лежат на полу камеры, как
обычно, а подвешиваются к ее потолку и стенам.
Помимо обычных камер, в которых находятся муравьи и расплод,
у некоторых видов имеются камеры для хранения запасов и камеры с отбросами.
Часто их строение не отличается от строения обычных камер Messor камеры
c запасами иногда имеют сферическую или близкую к ней форму. Камеры с отбросами
особенно характерны для Messor и Cataglyphis. У первых они заполнены
мякиной, а у вторых — хитиновыми остатками съеденных насекомых, в них обычно
встречается много неспециализированных мирмекофилов. Как у Messor,
так и у Cataglyphis обычно это большие плоские камеры часто в плане
неправильной формы, располагающиеся под самой поверхностью почвы.
У многих видов Cataglyphis, Proformion epinotalis
и Temnothorax semenovi гнездо представляет собой вертикальный ход (ствол),
с которым соеденены камеры. Иногда ствол ее соединяется непосредственно с
входом, а открывается в поверхностную галереи. В некоторых случаях эта галерея
может ветвиться, иногда расширяется и имеет вид длинной плоской камеры (C.oxiana),
иногда с ней соединяются несколько камер.
Наиболее распространенный тип подземных гнезд — гнезда с развитой
системой поверхностных галерей и камер на глубине до 20-40 см и одним или
несколькими стволами. Такие гнезда характерны для Messor, Tetramorium
и т.д. Гнезда молодых семей всегда имеют один ствол. Все стволы таких односекционных







гнезд имеют в принципе одинаковое строение и несут сходные функции, поскольку
в камерах каждого ствола имеются и расплод, и (у зерноядных видов) запасы
семян.
У Messor variabilis гнезда татже односекционные, но разные
стволы гнезда несут различные функции: одни из стволов соединены с входом
и глубинной горизонтальной системой галерей и камер, в которых хранятся запасы
семян, тогда как ствол, в котором находится расплод, не имеет самостоятельного
входа, а соединяется с глубинной горизонтальной системой. У этого вида вокруг
входа в гнездо сооружается надземная постройка, похожая на вулкан с глубоким
кратером. В ней располагаются камеры с остатками пищи.
При увеличения размеров семьи может наступить момент, когда
гнездо, даже имеющее несколько стволов, не может вместить все население. В
этом случае муравьи рядом с основным гнездом строят новые — второе, третье
и т.д. Если эти гнезда соединены подземными галереями, система носит название
многосекционного гнезда, а отдельные гнезда — секций. Если же сообщение между
отдельными гнездами происходит по поверхности почвы, по временным или постоянным
дорогам, мы говорим о поликалической семье. Многосекционные гнезда в простейшем
случае представляют собой линейную цепочку секций, соединенных подземными
галереями. Такой тип гнезда часто наблюдавтея у Pheidole pallidula
и др. У олигогинных видов (неапример Pl. pallescens) самки более или
менее равномерно распределены по секциям. В случае моногонии самка располагается
в одной из секций, что является предпосылкой для возникновения функциональной
дифференциации последних.
Поликалия характерна для представителей рода Grematogaster.
Гнезда с наружными постройками (холмиками), весьма характерные
для многих муравьев лесной зоны.
Систематический анализ полужесткокрылых Джейрахской и Таргимской аридных котловин Северо-Восточной части Большого Ї Кавказа
По результатам обработки материалов собственных сборов и по литературным источникам на территории Джейрахской и Таргимской аридных котловин Северо-Восточной части Большого Кавказа было зарегистрировано 157 видов полужесткокрылых, из которых 95 видов указываются впервые для района исследования. .. Распределение видов по семействам представлено в таблице 1. Как видно из таблицы фауна полужесткокрылых Джейрахской и Таргимской аридных котловин Северо-Восточной части Большого Кавказа состоит из 20 семейств, включающих 109 родов. Наибольшим числом видов представлены семейства: MIRIDAE (более 35%), LYGAEIDAE (16,56 %) и PENTATOMIDAE (14,65%). Такое распределение видов по таксонам характерно для умеренных широт.
Наиболее богаты видами следующие роды (в скобках число видов): Orthops (4 ), Stenodema (4 ), Carpocoris (4 ), Heterogaster (3), Megalonotus (3), Nysius (3), Adelphocoris (3), Lygus (3), Eurygaster (3).
В списке приведены все виды полужесткокрылых, указанные для региона в литературе (Абдурахманов, Драполюк, 1990;) и собранные автором Гадаборшевой М.А. в 2000-2005 гг. на данной территории при участии соискателя Гелисхановой СБ. Для каждого вида приводится ссылки на литературные источники, в которых они впервые указывались для Большого Кавказа, места и дата сборов, кратко излагаются стациально-биотопическая приуроченность, принадлежность к экологической группе, пищевая специализация (для некоторых видов и вредоносное значение), жизненный цикл и распространение вида с указанием типа ареала. Данные о распространении видов приведены с согласно сведений многих авторов (Драполюк, 1989; Бондаренков, 1949; Кириченко, Винокуров, 1978; Кириченко, Ячевский, 1960; В.Г.Пучков, 1961, 1970, 1971, 1975, 1977, 1978; П. В. Пучков, 1978, 1982, 1991; В.Г.Пучков, В. П. Пучков, 1979; Талыцкий, Пучков, 1966; Зайцева,1968, 1977, 1984, 1981,1997, 1998; Гидаятов, 1967, 1971, 1970, 1972, 1975, 1981; Заводчикова, 1983; Быков, 1972; Буров, 1963; Кержнер, 1962, 1964а, 1970, 1972, 1977, 1979, 1981, 1988; Элов, Кержнер, 1977; Яковлев, 1905; Элов, 1976; Коровино-Положенцева, 1959; Писарев, Мещеряков, Попов, 1969; Кириченко, 1918, 1938, 1951, 1952, 1964; Держанский, 1989, 1994, 1990, 2000; Гидаятов, Атакишиева, 1974, 1981; Гидаятов, Драполюк, Атакишиева, 1980; Чернова, 1978, 1979; Канюкова, 1973, 1982, 1989; Пучкова, 1968; Скляр, 1971; Акрамовская, 1959; Асанова, 1964, 1971, 1975, 1979; Винокуров, 1979, 1981, 1982, 1990; Винокуров, Конюкова, 1995; Голуб, 1974, 1976, 1977, 1978, 1989, 1999; Нейморовец, 2003; 2004; Oshanin, 1910, 1912; Breddin, 1907; Distant, 1910;Wagner, 1974, 1975a, 19756; Stichel, 1958; 1959; Linnavuori, 1960, Kelton, 1963; Slater, 1964; Asanova, Kerzhner, 1969; Josifor, Kerzhner, 1978; Aukema, Rieger, 1995, 1996, 1999, 2001; Derzhansky, 1997; Henry, Froeschner, 1988; Kanyukova, 1997; Konstantinov, 1997; Lukashuk, 1997; Moulet, 1995; Neimorovets, 2001; Aglymzyanov, 1994).





1. Elasmostethus interstinctus (Linnaeus, 1758) Лит.: Кириченко (1918). Мат.: Таргимская котловина, разв. Н. Озиг, 6 VI2004,14 VII2004. Экол. и биол.: тамно-дендробионт (по данным Пучкова (1965) и И.М. Кержнера (1964) живет на видах p. Betula, p. Alnus и буке); мезофил; полифитофаг (по данным Р.Б. Асановой и Б.В.Искакова (1977) обнаружен на жимолости карагаче, а также на лиственных деревьях и кустарниках; наблюдается каннибализм); моновольтинный; зимуют имаго. Распр.: Европа (кроме южной), Кавказ и Закавказье, Ю. Сибирь, Ср. Азия, Д. Восток, Канада, США — голарктический циркумбореальный
2. Elasmucha grisea (Fieber, 1861); на территории Джейрахской котловины вид представлен подвидом Е. g. grisea. Лит.: Пучков (1967), Абдурахманов и Драполюк (1990). Мат.: Джейрахская котловина (1990). Экол. и биол.: дендробионт (по данным И.М. Кержнера (1972) живет на березе и ольхе; Д. А. Гидаятовым (1982) отмечен в Азербайджане на дубе и буке; а также на тополе и иве); мезофил; широкий олигофитофаг (Betula, Alnus); моновольтинный; зимуют имаго. Распр.: Европа (кроме южной), Сибирь, Ср. Азия, Д. Восток транспалеарктический бореальный.
3. Elasmucha ferrugata (Fabricius, 1787) Лит.: Кириченко (1918). Мат.: Таргимская котловина, подножие г. Лейми-Лом, 27 VIII 2005. Экол. и биол.: дендробионт (на черемухе, боярышнике); мезофил; широкий олигофитофаг (на ягодных кустарниках: Ribes, Vaccinium, Grossularia и т.д.); возможно, моновольтинный; зимуют имаго Распр.: Европа (кроме южной), Сибирь, Монголия, Ср. Азия, Д. Восток -транспалеарктический бореальный.
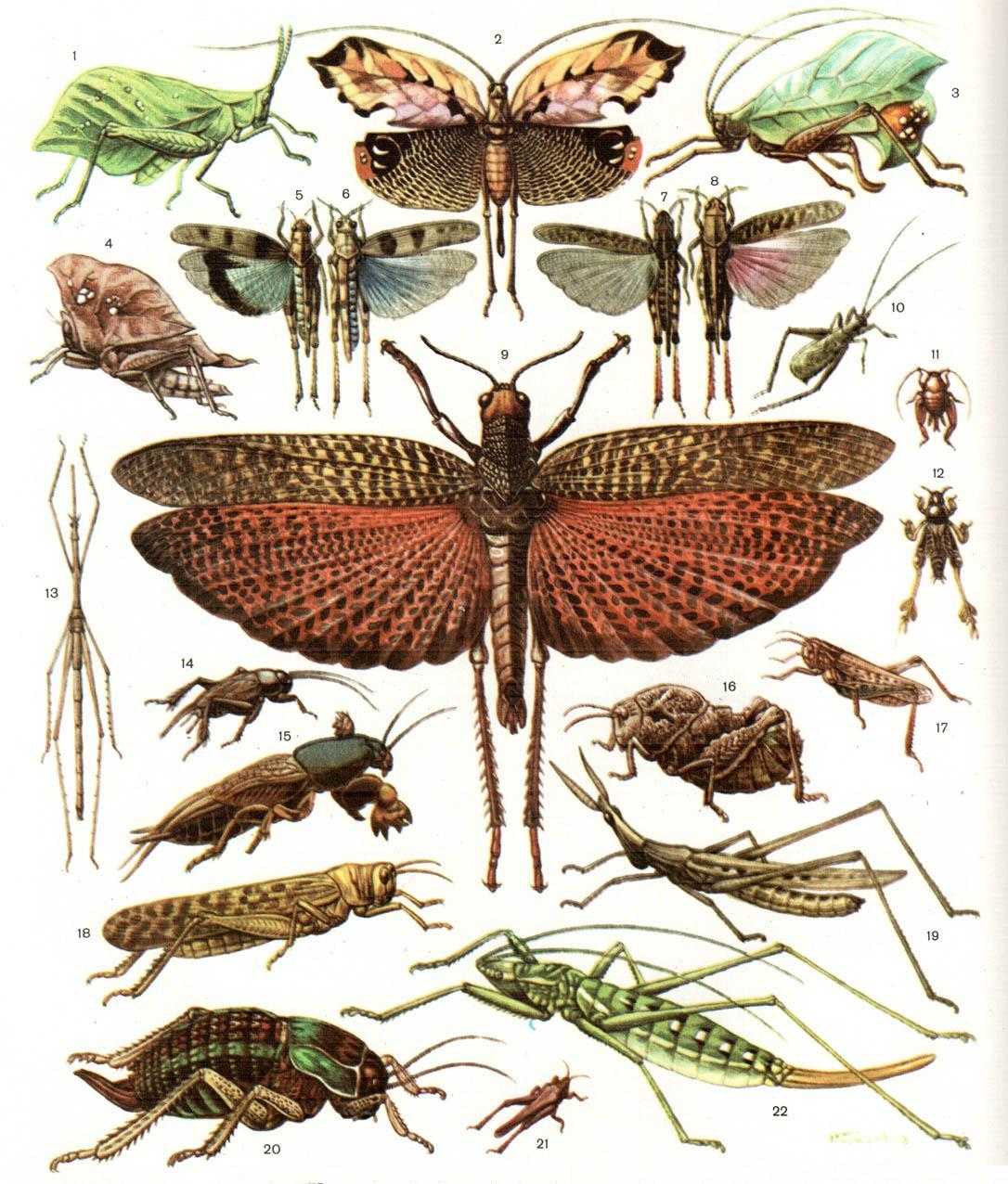
Хортобионты
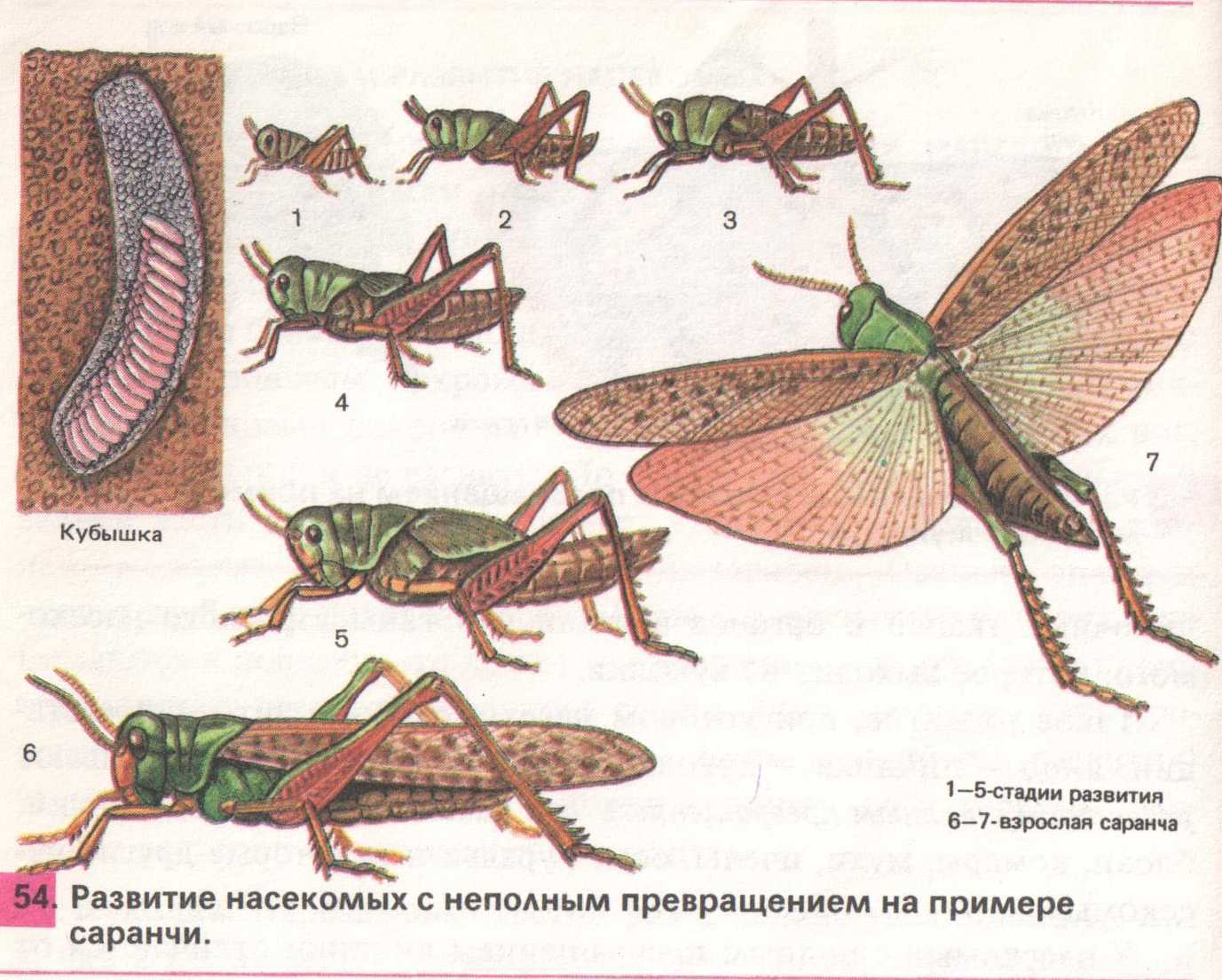
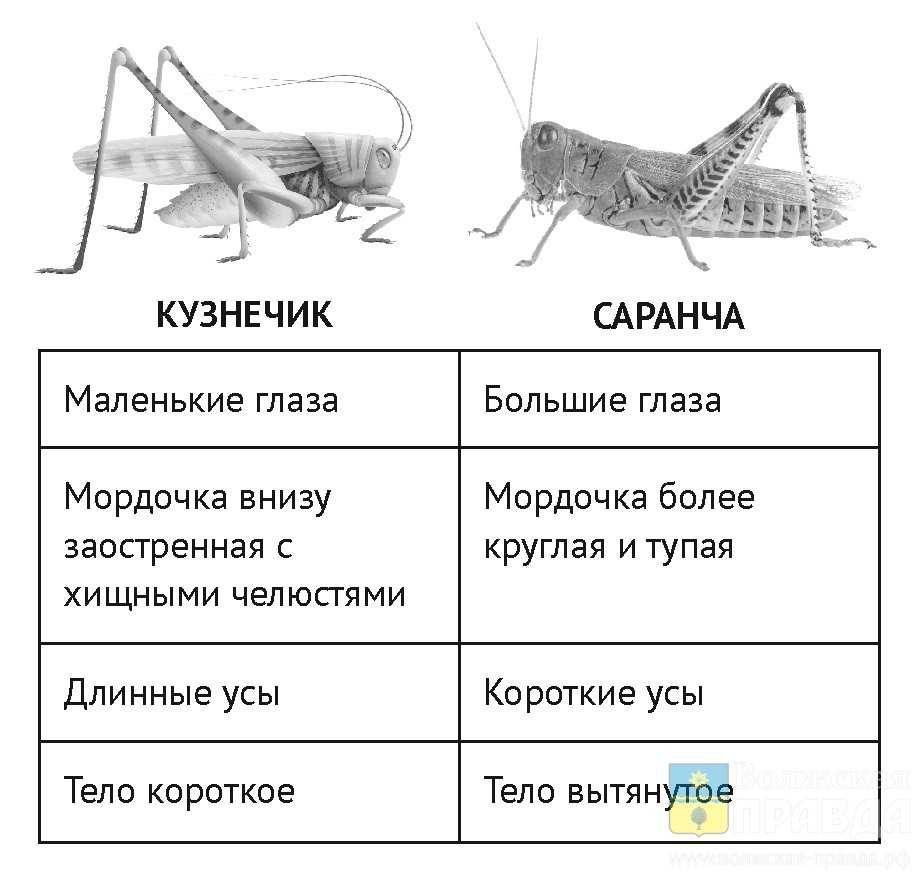
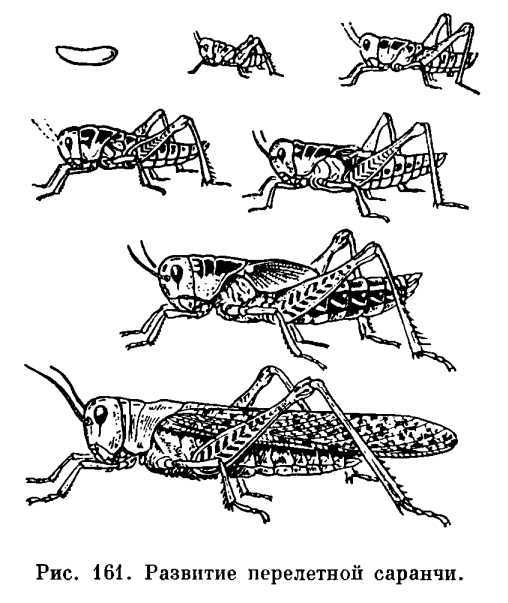
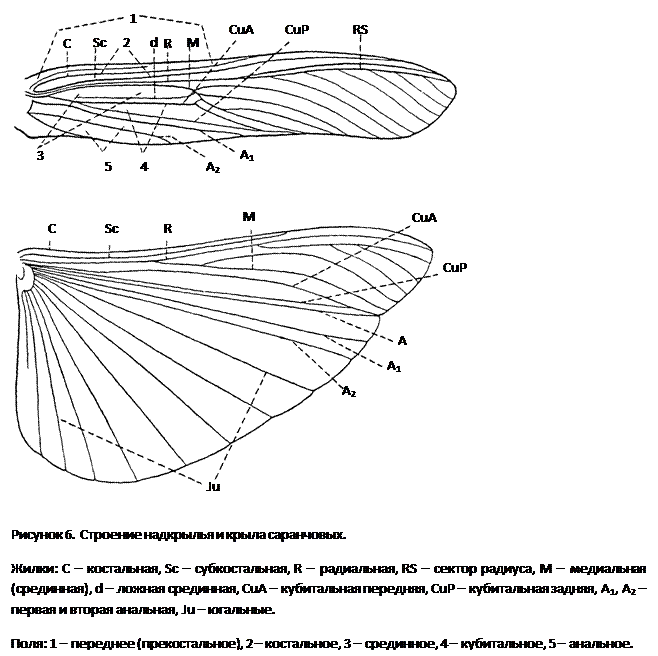
У них стройное удлиненное тело, нередко со скошенным лбом; соотношение между шириной и высотой тела в наиболее широкой его части (индекс ш/в) всегда ниже единицы. В некоторых случаях тело может быть настолько сильно удлинено и к тому же лишено крыльев, что приобретает палочковидную форму, как это имеет место у южноамериканской палочковидной кобылки (Cephalocoema lineata).
Покровы тела хортобионтов гладкие, лишены грубой скульптуры и окрашены под цвет зеленой или сухой травы, часто с продольными полосками вдоль тела. Крылья обычно бесцветные, реже окрашены в бледные розоватые или зеленоватые тона.
Хортобионты
Среди хортобионтов многие виды являются вредителями сельского хозяйства. Некоторые хортобионты предпочитают питаться не злаками, а широколиственными травянистыми растениями; их так и называют травоядными хортобионтами.
Сюда относятся виды бескрылых кобылок конофим (Соnophyma, табл. 30, 4), широко распространенных в горах Средней Азии, и целый ряд других форм. У травоядных хортобионтов туловище более коренастое, а голова с отвесным лбом.
Характерно, что у них верхняя челюсть приспособлена к перетиранию и пережевыванию мягких листьев. В ряде случаев встречаются виды, которые хотя и обитают в толще травы, но не избегают и открытых пространств на поверхности почвы, как это делают некоторые крестовички (Dociostaurus).
Такие саранчовые называются факультативными хортобионтами. По своему поведению и форме тела они являются переходными к геофилам. В отличие от хортобионтов, связанных с травянистой растительностью, тамнобионты (табл. 30, 5, 6) живут на кустарниках и деревьях.
Приспособления к такому образу жизни прежде всего выражаются в специфичной структуре конечностей: у них наблюдается асимметрия в вооружении голеней задних ног, на внутренней стороне которых шипы длиннее шипов наружного ряда; асимметричными могут быть и находящиеся на лапках коготки, между которыми находится сильно развитая присоска.
Питание
Все виды муравьев, обитающих в одном биогеоценозе, образуют
коадаптивный комплекс (Длуccкий, 1981), члены которого определенным образом
делят между собой пищевые ресурсы биогеоценоза. Во влажных местообитаниях,
где более или менее выражена ярусность биогеоценозов, муравьи делят среду,
главным образом, по вертикали. Дендробионты (некоторые Camponotus, Crematogaster
и Leptothorax ) добывают пишу в кронах и на стволах деревьев, хортобионты
— в травостое, герпетобионты ( Tetramorium, Aphaenogaster, Formica
и т.д.) — на поверхности почвы, стратобионты (Myrmica, Ponera, Leptothorax
и др.) в подстилке, геобинты (Lasius flavus) — в почве. Питание этих
муравьев довольно сходно. Вот они являются зоонекрофагами, т.е. питаются малоподвижными
насекомыми или их трупами, а также посещают колонии тлей, от которых они получают
падь.
Можно выделять 3 главных трофических группы:
1. Карпофаги, или муравьи, питающиеся семенами растений. К этой
группе относятся муравьи-жнецы (Messor) и мелкие карпофаги (Tetramorium,
Pheidole, Monomorium и др.).
2. Дневные зоонекрофаги. В эту группу входят представителя родов
Cataglyphis, Proformica, Formica и др. Источником белка для них являются,
главным образом, трупы насекомых. В наибольшей степени это характерно для
крупных Cataglyphis. Большинство видов этой группы получает углеводную
пищу, собирая экскременты тлей, цикад либо. Многие муравья не только эксплуатируют,
но и охраняют колонии тлей от хищников и паразитов. Все Formicinae
и Dolichoderinae в той или иной степени способны создавать запасы жидкой
пищи, содержащей большое количество сахаров.
3. Ночные хищники. В эту группу входят представителя подрода




Tanaemyrmex рода Camponotus. В принципе характер питания Tanaemyrmex
не отличается от такового у других герпетобионтов-зоонекрофагов, они также
собирают трупы насекомых, охотятся на малоподвижных беспозвоночных на поверхности
почвы или собирают падь тлей. Однако эти насекомые начинают охоту после захода
солнца и эаканчивают ее утром.
Большенство зоофагов и зоонекрофагов имеют очень широкий спектр
питания, однако у части видов может наблюдаться определенная специализация.