
Протеины мембраны
Протеины мембраны — это интегральные белки, которые встречаются в мембранах всех живых организмов. Они играют важную роль в поддержании структуры мембраны и выполнении различных функций.
Протеины мембраны характеризуются своей внутренней структурой, аминокислотным составом и функциональными свойствами. Они способны взаимодействовать с другими молекулами, включая другие протеины, липиды и углеводы.
Протеины мембраны играют важную роль в регуляции множества биологических процессов, включая транспорт веществ через мембраны, клеточное распознавание, общение между клетками и многие другие функции. Понимание структуры и функций протеинов мембраны является важным для разработки новых лекарственных препаратов и диагностических методов.
- Транспортные протеины перемещают различные вещества через мембрану, что позволяет клетке получать необходимые питательные вещества и избавляться от отходов.
- Рецепторные протеины распознают молекулы сигнальных веществ, таких как гормоны, и инициируют цепочку внутриклеточных реакций.
- Ферменты являются катализаторами реакций в мембране и участвуют в многих биохимических процессах.
Протеины мембраны необходимы для правильной структуры и функции мембраны, а также для выполнения множества жизненно важных функций в клетке. Их изучение является основой для понимания биологических процессов и разработки новых методов лечения многих заболеваний.
Типы межклеточных соединений[]
Плазмодесмы
Микроскопические цитоплазматические мостики, соединяющие соседние клетки растений. Основная статья: Плазмодесмы
Простое межклеточное соединение
При простом межклеточном соединении оболочки клеток сближены на расстояние 15 – 20 нм. Это соединение занимает наиболее обширные участки соприкасающихся клеток. Посредством простых соединений осуществляется слабая механическая связь, не препятствующая транспорту веществ в межклеточных пространствах. Разновидностью простого соединения является контакт типа «замок», когда билипидные мембраны соседних клеток вместе с участком цитоплазмы вдавливаются друг в друга, чем достигается большая поверхность соприкосновения и более прочная механическая связь.
Плотное соединение (запирающая зона)
В плотном соединении клеточные мембраны максимально сближены, здесь фактически происходит их слияние. Роль плотного соединения заключается в механическом сцеплении клеток и препятствии транспорту веществ по межклеточным пространствам. Эта область непроницаема для макромолекул и ионов, она ограждает межклеточные щели от внешней среды. Плотные соединения обычно образуются между эпителиальными клетками в тех органах (желудке, кишечнике и пр.), где эпителий ограничивает содержимое этих органов (желудочный сок, кишечный сок). В этих участках плотные контакты охватывают по периметру каждую клетку, межмембранные пространства отсутствуют, а соседние клеточные оболочки слиты в одну. Если же плотное сцепление происходит на ограниченном участке, то образуется пятно слипания (десмосома).Частными случаями плотного соединения являются зоны замыкания и слипания.
Зона замыкания
В зоне замыкания две соседние мембраны сливаются своими наружными слоями, эта зона непроницаема для макромолекул и ионов.
Зона слипания (промежуточный контакт)
В зоне слипания мембраны разделены щелью в 10-20 нм, заполненной плотным веществом (белковой природы).
Десмосома (пятно сцепления, липкое соединение)
Основная статья: Десмосомы
Десмосома представляет собой небольшую площадку, иногда слоистого вида, диаметром до 0,5 мкм. Их функциональная роль заключается главным образом в механической связи между клетками. Существуют 3 типа десмосом – точечные, опоясывающие и полудесмосомы. Десмосомой называется образованное клетками соединение, прочно склеивающее клетки. Если они образуются между клетками и внеклеточным матриксом, то они называются полудесмосомами. Количество десмосом на одной клетке может достигать 2000. Такие контакты встречаются между клетками, которые могут подвергаться трению и другим механическим воздействиям (эпителиальные клетки, клетки сердечной мышцы). Со стороны цитоплазмы к десмосомам прикрепляются промежуточные филаменты, которые формируют остов цитоплазмы, обладающий большой прочностью на разрыв. Таким образом, через десмосомы промежуточные филаменты соседних клеток объединяются в непрерывную сеть по всей ткани. Тип промежуточных филаментов зависит от типа клеток: в большинстве эпителиальных клеток они кератиновые, а в клетках сердечной мышцы – десминовые.
Нексус (щелевой контакт)
Нексус представляет собой ограниченный участок контакта двух клеточных мембран диаметром 0,5 – 3 мкм с расстоянием между мембранами 2-3 нм. Обе эти мембраны пронизаны белковыми молекулами коннексонами, содержащими гидрофильные каналы. Через эти каналы осуществляется обмен ионами и микромолекулами соседних клеток. Поэтому нексусы называют также проводящими соединениями. Их функциональная роль заключается в переносе ионов и мелких молекул от клетки к клетке, минуя межклеточное пространство. Этот тип соединения встречается во всех группах тканей.
Синапсы являются особыми формами межклеточных соединений. Они характерны для нервной ткани и встречаются между нейронами (межнейронные синапсы) или между нейроном и клеткой-мишенью (нервно-мышечные синапсы и пр.). Синапсы – участки контакта двух клеток, специализированных для односторонней передачи возбуждения или торможения от одной клетки к другой. Их функция – именно передача нервного импульса с нейрона на другую нервную клетку или клетку-мишень.
Эндоцитоз
Эндоцитоз – это процесс, при котором клетка захватывает материал из внешней среды, образуя внутриклеточные везикулы, покрытые мембраной. Процесс осуществляется с помощью специальных белков, находящихся на поверхности клетки.
Существует два вида эндоцитоза:
-
-
- Фагоцитоз – процесс захвата крупных частиц, таких как бактерии или мертвые клетки, клетками с помощью псевдоподий.
- Пиноцитоз – процесс захвата питательных веществ в жидкой среде клеткой.
-
В ходе эндоцитоза, клетка создает внутри себя мембранные пузырьки, которые поглощают частицы из внешнего окружения. После захвата и переноса внутрь клетки, эти частицы могут быть переработаны или использованы клеткой для своих потребностей.
Эндоцитоз является важным механизмом выживания и функционирования клеток, особенно для клеток-фагоцитов. Кроме того, благодаря эндоцитозу, клетки регулируют содержание растворов внутри себя, поддерживая стабильность своей внутренней среды.
Проводящие ткани (ксилема, флоэма)
Ксилема – сложная ткань, то есть состоит из клеток разной морфологии. В состав ксилемы одновременно входят и проводящие, и механические, и запасающие элементы.
Ксилема проводит воду с растворенными в ней минеральными веществами от корней по всему остальному телу растения. Таким образом, по ксилеме в основном осуществляется восходящий ток. Проводящие элементы ксилемы – это сосуды и трахеиды. Следует помнить, что ксилема голосеменных растений лишена сосудов. Трахеида образуется из клетки удлиненной формы, ее клеточная стенка утолщается и лигнифицируется, то есть одревесневает. Протопласт при этом отмирает и в результате получается мелкий капилляр, по которому может транспортироваться вода. Прочные клеточные стенки предохраняют просвет капилляра от схлопывания. От трахеиды к трахеиде вода транспортируется через специальные поры. Сосуд, по сути, является таким же капилляром, как и трахеида, но более длинным, широкопросветным и многоклеточным. Каждый сосуд состоит из отдельных клеток (члеников сосуда) с одревесневшей оболочкой и отмершим протопластом, между члениками сосуда формируются уже не поры, а перфорационные пластинки (то есть сквозные отверстия). Между сосудами, как и между трахеидами, есть поры, через которые также может транспортироваться вода. Кроме проводящих элементов, в состав ксилемы входят механические волокна – волокна либриформа. Это удлиненные клетки, похожие на трахеиды, однако их клеточные стенки очень сильно утолщены и лигнифицированы. Просвет таких капилляров слишком мал для осуществления транспорта воды, зато толстая и прочная клеточная стенка выполняет механическую функцию подобно склеренхиме. Ксилема в основном состоит из мертвых клеток, обычно небольшой процент живых клеток представлен древесинной паренхимой. Эти клетки в основном выполняют запасающую функцию.
Флоэма, как и ксилема, – это сложная ткань, которая состоит из разных клеток. В состав флоэмы входят проводящие механические и паренхимные (в том числе запасающие) элементы.
Флоэма транспортирует раствор питательных веществ, в основном это углеводы, образовавшиеся в результате фотосинтеза. Поскольку фотосинтез происходит преимущественно в листьях, а питательные вещества нужно доставлять во все части растения, в том числе и в корни, по флоэме преимущественно осуществляется нисходящий ток веществ. Проводящими элементами являются ситовидные клетки. Это живые клетки, они имеют вытянутую форму, а в их стенках формируются так называемые ситовидные поля. Ситовидное поле – это участок клеточной стенки, где близко друг к другу расположено множество плазмодесм. Через ситовидные поля происходит транспорт веществ от одной ситовидной клетки к другой. У покрытосеменных растений проводящими элементами флоэмы являются ситовидные трубки. Ситовидная трубка – это более длинная многоклеточная проводящая структура. Состоит она из одного ряда клеток, называемых члениками ситовидной трубки. В местах контакта члеников друг с другом формируются ситовидные пластинки – участки клеточной стенки, где расположено одно или несколько сближенных ситовидных полей. Вещества транспортируются по внутреннему содержимому живой клетки. Однако в ситовидных элементах деградируют многие органеллы, в том числе и ядро. Таким образом, ситовидная клетка и членик ситовидной трубки находятся в «полуживом» состоянии. При этом существуют специальные клетки, которые поддерживают ситовидные элементы в этом состоянии, обеспечивают и регулируют их жизнедеятельность. Такие клетки называются клетками-спутницами у члеников ситовидных трубок, а ситовидные клетки поддерживают специальные клетки Страсбургера. Кроме проводящих элементов во флоэме, как и в ксилеме, находятся паренхимные (запасающие) клетки, а также механические элементы (лубяные волокна). Волокна обычно представлены удлиненными клетками с толстой одревесневшей клеточной стенкой.
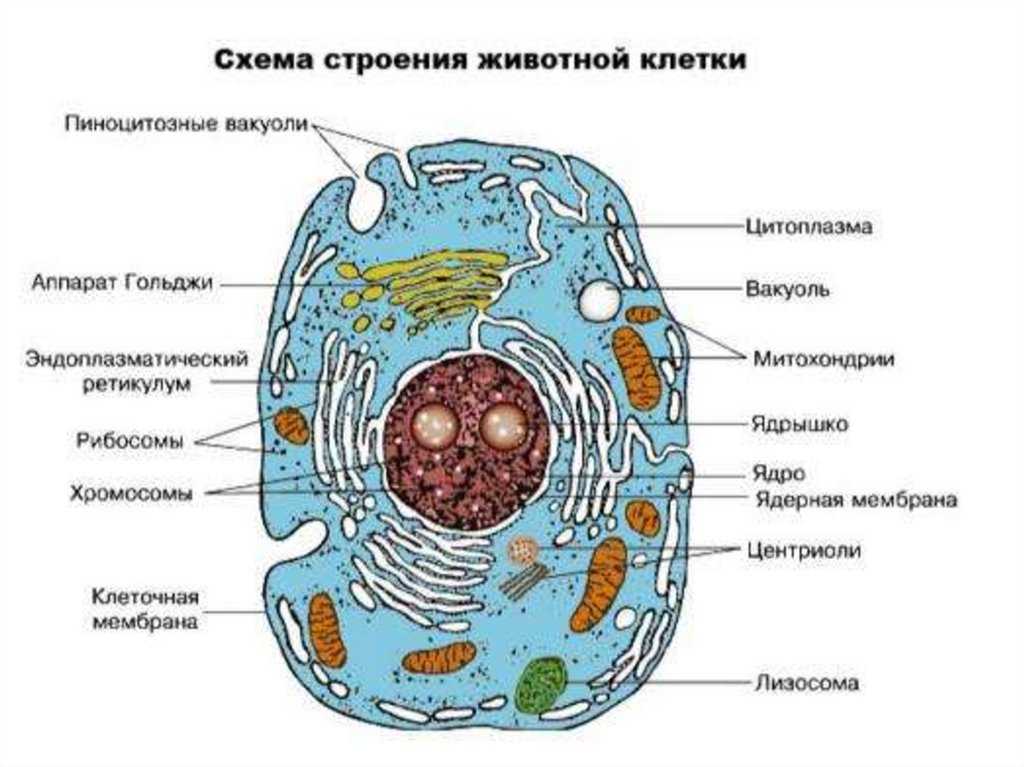


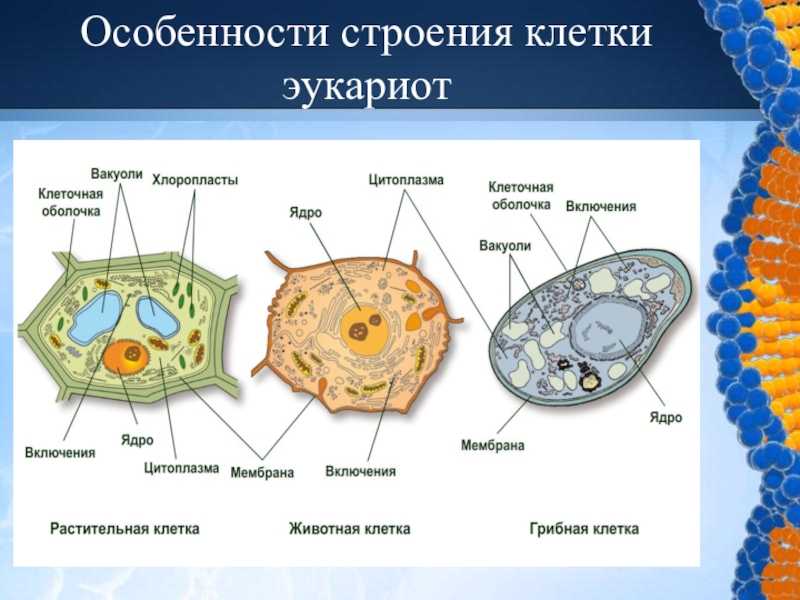
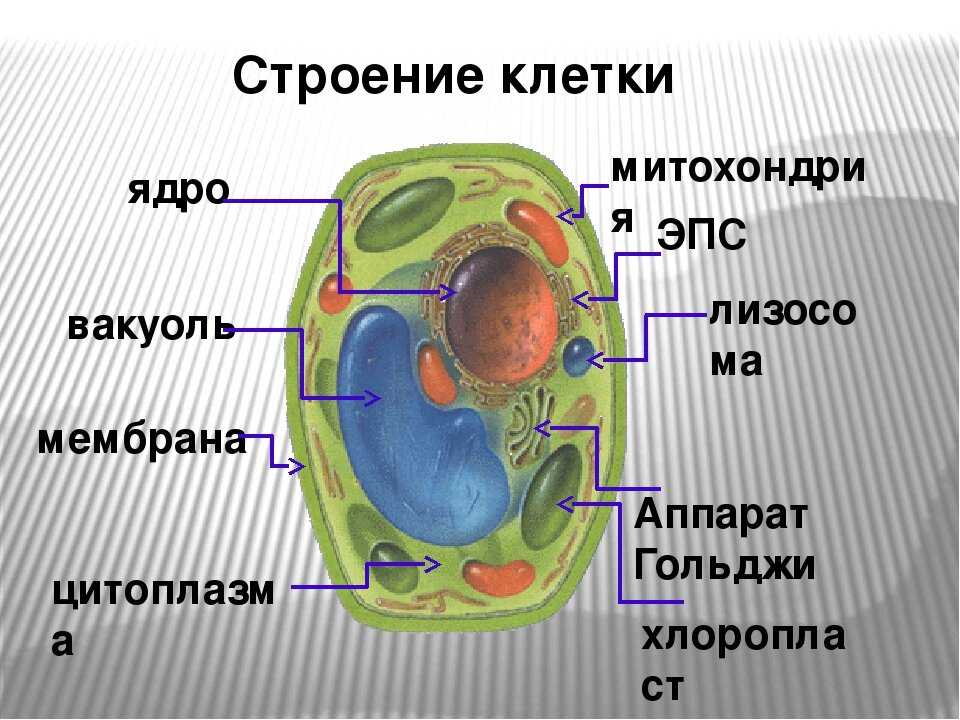
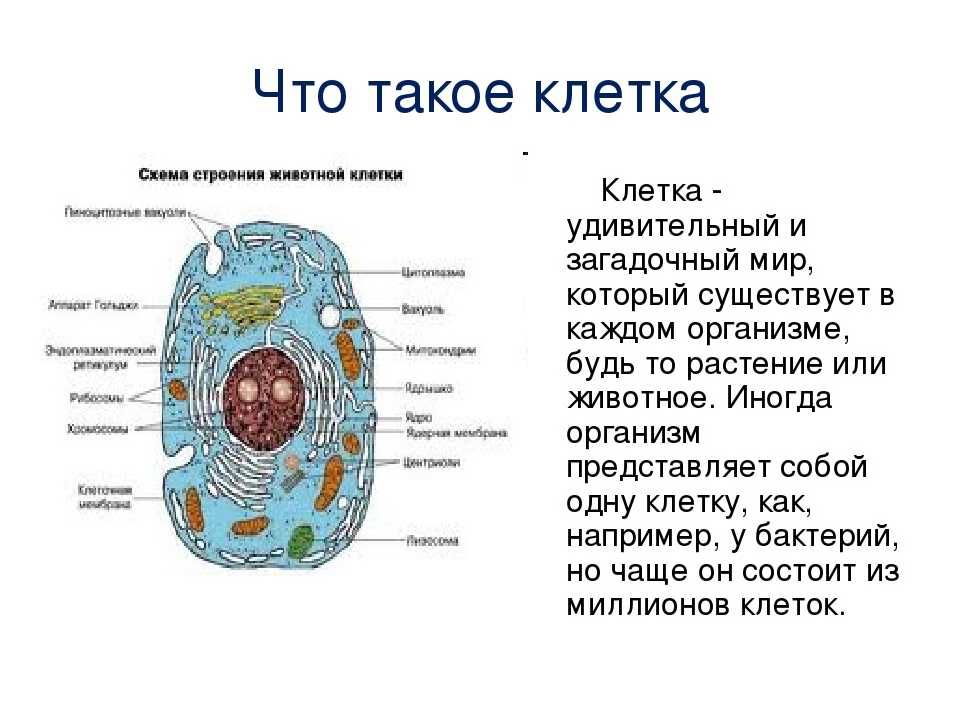
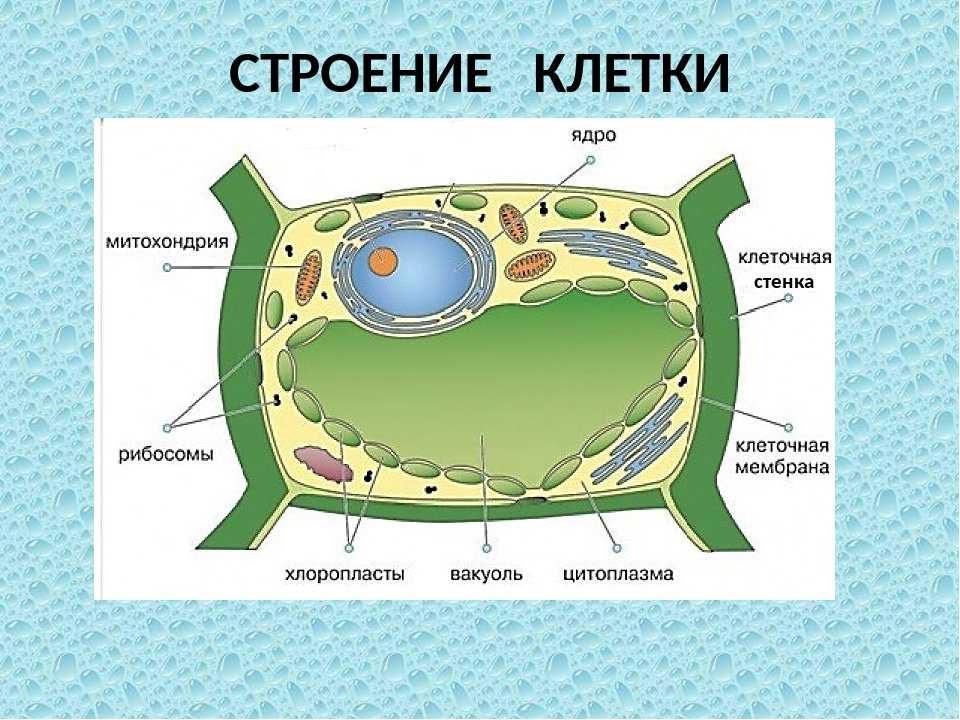

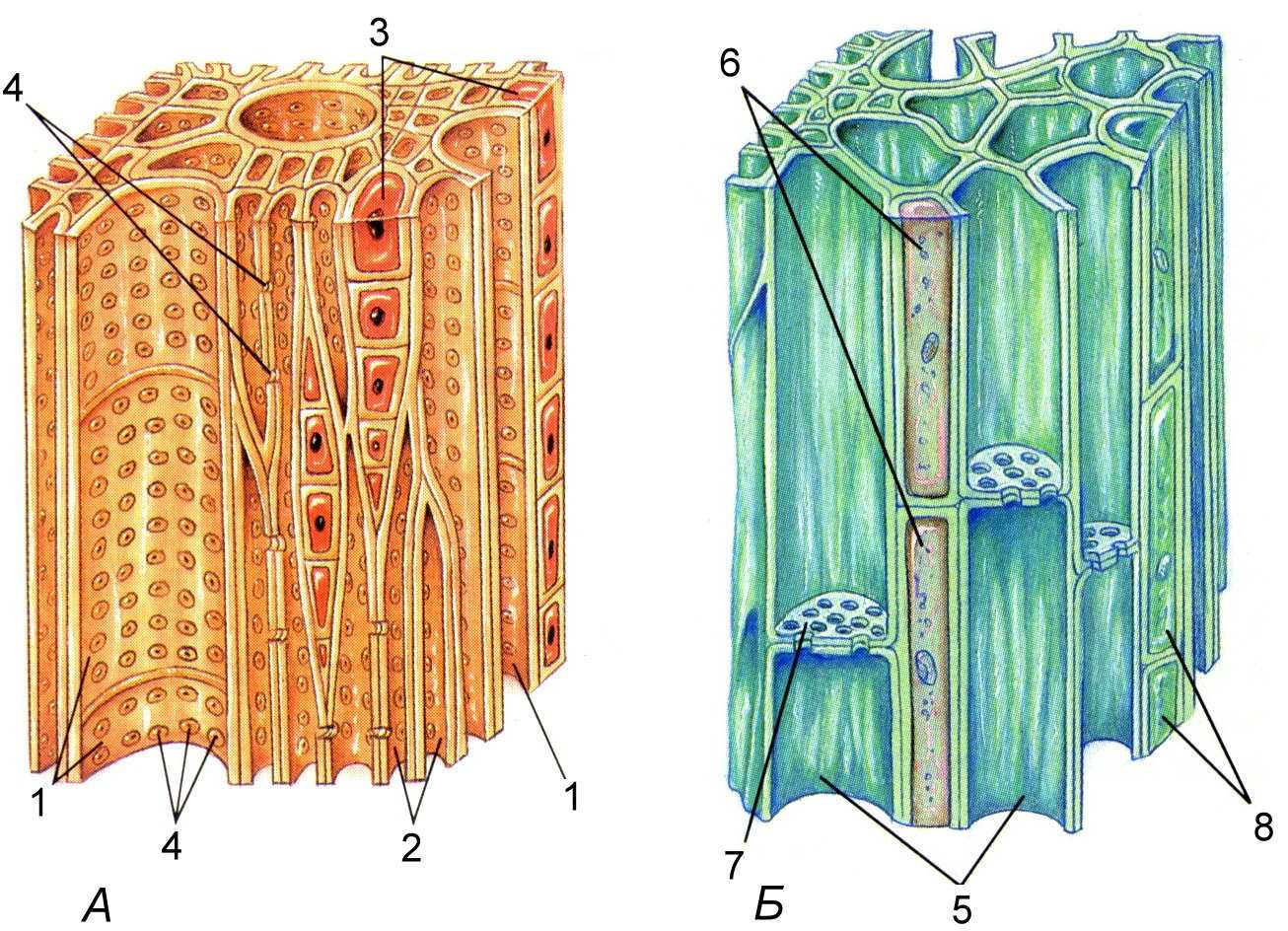
Рисунок: Проводящие ткани. А – ксилема; Б – флоэма. 1 – сосуды ксилемы; 2 – трахеиды; 3 – клетки древесной паренхимы; 4 – поры; 5 — ситовидные трубки; 6 – клетки – спутницы; 7 – ситовидные поля; 8 – клетки лубяной паренхимы.
Поглощающие ткани
Высшие растения поглощают воду с помощью специальных тканей. У мохообразных отсутствуют корни, и всасывание воды происходит всей поверхностью тела (например, с помощью гиалиновых клеток у сфагновых мхов) или с помощью ризоидов – длинных тонкостенных клеток. Сосудистые растения имеют корни, поверхность которых покрыта ризодермой (эпиблемой) – специализированной всасывающей тканью. Ризодерма гомологична эпидерме, то есть также формируется из одного внешнего слоя клеток, покрывающих орган. Однако ризодерма не является покровной тканью, поскольку практически не выполняет защитную функцию. Ее клетки тонкостенные и специализируются на поглощении воды и минеральных солей из почвы, поглощение при этом происходит избирательно и с затратой энергии. В ризодерме различают два типа клеток: трихобласты и атрихобласты. У трихобластов наружная часть клетки выпячивается и образует длинный вырост – корневой волосок, служащий для увеличения поверхности всасывания. Корневой волосок выделяет слизь, которая помогает растворять поглощать минеральные вещества из почвы. Атрихобласты не формируют корневых волосков, но также поглощают вещества своей поверхностью.
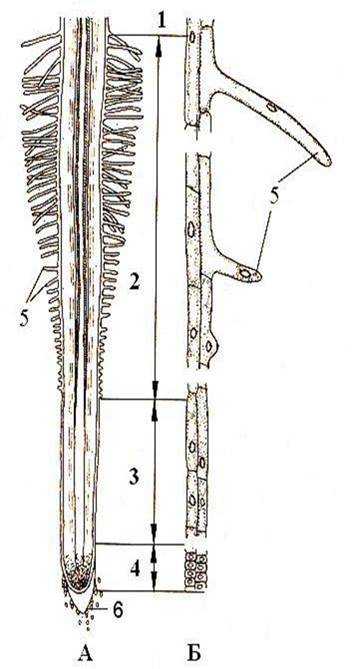
Рисунок: Ризодерма. А – Продольный разрез корня; Б – Клетки ризодермы. 1 – зона проведения; 2 – зона всасывания; 3 – зона роста; 4 – зона деления; 5 – корневые волоски; 6 – корневой чехлик.
У некоторых тропических эпифитных растений вместо ризодермы развивается веламен. Веламен гомологичен ризодерме, но в отличие от нее является многослойной тканью и состоит из отмерших клеток. Их клеточные стенки имеют спиральные утолщения, которые служат ребрами жесткости, сами клеточные стенки частично разрушаются, а внутреннее содержимое клеток отмирает. В результате получается структура наподобие губки, которая способна впитывать воду из влажного воздуха, тумана или осадков. Таким образом, веламен поглощает вещества пассивно и не избирательно. Направленный и избирательный транспорт воды дальше внутрь корня происходит при участии экзодермы, подстилающей веламен (как, впрочем, и любую ризодерму).
В многоклеточных организмах
В многоклеточном организме передача сигналов между клетками происходит либо через высвобождение во внеклеточное пространство, разделенное на паракрин. передача сигналов (на короткие расстояния) и эндокринные сигналы (на большие расстояния) или путем прямого контакта, известная как передача сигналов юкстакринов. аутокринная передача сигналов является особый случай паракринной передачи сигналов, когда секретирующая клетка имеет способность отвечать на секретируемую сигнальную молекулу. Синаптическая передача сигналов — это особый случай паракринной передачи сигналов (для химических синапсов ) или юкстакринных сигналов ( для электрических синапсов ) между нейронами и клетками-мишенями. Сигнальные молекулы взаимодействуют с клеткой-мишенью в качестве лиганда к рецепторам клеточной поверхности и / или проникая в клетку через ее мембрану или эндоцитоз для внутрикринной передачи сигналов. Обычно это приводит к активации вторичных мессенджеров, что приводит к различным физиологическим эффектам.
Конкретная молекула обычно используется в различных режимах передачи сигналов, и поэтому классификация по способам передачи сигналов невозможна. По крайней мере, три важных класса сигнальных молекул широко известны, хотя и не являются исчерпывающими и с неточными границами, поскольку такое членство не является исключающим и зависит от контекста:
- Гормоны являются основными сигнальными молекулами эндокринная система, хотя они часто регулируют секрецию друг друга посредством локальной передачи сигналов (например, островок клеток Лангерганса ), и большинство из них также экспрессируются в тканях для местных целей (например, ангиотензин ) или в противном случае структурно родственные молекулы (например, PTHrP ).
- нейротрансмиттеры являются сигнальными молекулами нервной системы, включая также нейропептиды и нейромодуляторы. Нейротрансмиттеры, такие как катехоламины, также секретируются эндокринной системой в системный кровоток.
- Цитокины являются сигнальными молекулами иммунной системы, с первичной паракринной или юкстакринной ролью, хотя во время значительных иммунных реакций они могут иметь e сильное присутствие в кровотоке с системным действием (изменение метаболизма железа или температуры тела ). Факторы роста могут рассматриваться как цитокины или другой класс.
Сигнальные молекулы могут принадлежать к нескольким химическим классам: липиды, фосфолипиды, амино кислоты, моноамины, белки, гликопротеины или газы. Сигнальные молекулы, связывающие поверхностные рецепторы, обычно большие и гидрофильные (например, TRH, вазопрессин, ацетилхолин ), тогда как те, что входят в клетку, обычно небольшие и гидрофобные (например, глюкокортикоиды, гормоны щитовидной железы, холекальциферол, ретиноевая кислота ), но важные исключения из оба многочисленны, и одна и та же молекула может действовать как через поверхностный рецептор, так и внутрикринным образом, оказывая различные эффекты. При внутрикринной передаче сигналов, оказавшись внутри клетки, сигнальная молекула может связываться с внутриклеточными рецепторами, другими элементами или стимулировать активность фермента (например, газы). Внутрикринное действие пептидных гормонов остается предметом споров.
продуцируется в небольших количествах некоторыми клетками человеческого тела и выполняет ряд функций биологической передачи сигналов. В настоящее время известно, что только два других таких газа действуют как сигнальные молекулы в организме человека: оксид азота и .
У растений
Передача сигналов в растениях осуществляется через растительные гормоны, фитохромы, криптохромы и т. д.
Важными семействами растительных гормонов являются ауксин, цитокинин, гиббереллин, этилин, жасмоновая кислота, салициловая кислота, стриголактоны, полиамины, оксид азота, пептидные гормоны и т. Д. Также сообщалось о транслокации РНК
Вычислительные модели
Недавние подходы к лучшему пониманию элементов перекрестного взаимодействия путей, комплексному связыванию лигандному- рецептор и динамике сигнальной сети способ использования подходов системной биологии. Вычислительные модели часто нацелены на сбор информации из опубликованной литературы для создания связного набора компонентов сигнализации и связанных с ними взаимодействий. Развитие вычислительных моделей позволяет более глубоко исследовать сигнальные пути клеток на глобальном уровне, манипулирующие различные переменные и систематически оценивая результирующий ответ. Использование аналитических моделей для изучения передачи сигналов широко применяется в областях фармакологии и открытия лекарств для оценки взаимодействий рецептор-лиганд и фармакокинетики, а также потока метаболитов в больших сетях. Обычно применяемая стратегия для моделирования клеточных сигнальных механизмов заключается в использовании моделей обыкновенного дифференциального уравнения (ODE) путем выражения зависящей от времени концентрации сигнальной молекулы как функции других молекул ниже и / или выше в пределах путь. Модели ODE уже применялись для динамического анализа сигнальных путей митоген-активированной протеинкиназы, рецептора эстрогена альфа и MTOR среди множества других.
Вентиляционные ткани (аэренхима)
Аэренхима – это вентиляционная ткань или ткань проветривания. Главную функцию аэренхимы выполняют крупные межклетники, по которым и циркулирует воздух. Воздух необходим высшим растениям как для дыхания, так и для процессов фотосинтеза. Наличие аэренхимы характерно для водных или околоводных высших растений. Воздух, находящийся в системе полостей аэренхимы, не только вентилирует все части растения (в особенности подводные), но и придает им плавучесть, как, например, листьям кувшинки.
Аэренхима обычно имеет вид системы полостей с однослойными стенками. Клетки, слагающие стенки полостей могут иметь вытянутую форму или же могут быть шарообразной формы. Сами полости при этом в некоторых местах имеют тонкие пленчатые перегородки из одного ряда мелких клеток. Клетки этих перегородок имеют звездчатую форму, таким образом, между «лучей» данных клеток остаются мелкие отверстия в пленке (межклетники). Данные перегородки не мешают выполнять вентиляционную функцию аэренхиме, пропуская воздух через эти мелкие отверстия. Однако, если произойдет повреждение и полость начнет заполняться водой, то такая перегородка не попустит капельно-жидкую воду, поскольку поверхностное натяжение жидкости не позволит ей пройти сквозь мелкие отверстия. Такая аэренхима встречается у кувшинки, ириса, рдеста и т.д.
В другом случае аэренхима может быть целиком представлена только звездчатыми клетками. Такие клетки формируют трехмерную рыхлую ткань, похожую по консистенции на вату. Между «лучей» этих клеток также формируется одно большое общее межклеточное пространство, по которому циркулирует воздух. Такой тип аэренхимы характерен для ситников, осок, некоторых злаков и т.д. Также рыхлая аэренхима, многократно преломляя свет, придает белый цвет лепесткам некоторых растений.
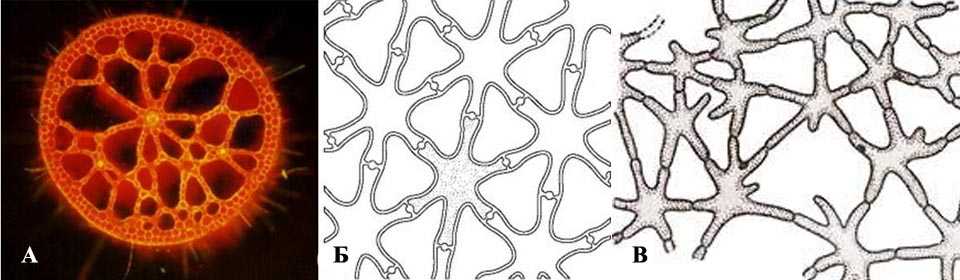
Рисунок: Аэренхима. А – аэренхима на поперечном срезе стебля; Б – клетки пленчатой перегородки, разделяющей полости аэренхимы; В – аэренхима из трехмерно расположенных звездчатых клеток.
Вентиляционная ткань выполняет свою функцию за счет многочисленных увеличенных межклетников. Стоит помнить, что межклетники по типу происхождения делятся на три типа. Схизогенные межклетники образовались в результате простого расхождения клеток в пространстве. Лизигенные полости формируются в результате деградации (лизиса) некоторых клеток. Крупные рексигенные полости являются результатом механического разрыва тканей, например, в центре черешков или стеблей некоторых растений.
# Анатомия растений
# 10 класс
# 11 класс
Холестерин в мембране
Холестерин — это немного жироподобное вещество, которое присутствует в мембранах всех живых клеток. Он играет важную роль в структуре и функционировании мембраны.
Интересно, что у разных организмов и типов клеток уровень холестерина в мембране может сильно варьироваться. Например, в клетках печени уровень холестерина в мембране выше, чем в других клетках. Также у некоторых бактерий мембрана содержит намного меньше холестерина, чем у животных клеток.
Холестерин — важный компонент мембраны, который обладает многими функциями. Его уровень и распределение в мембране могут быть различными у разных организмов и клеток. Однако он всегда играет ключевую роль в поддержании устойчивости мембраны и ее свойств.
Механические ткани
Существует две специализированные механические ткани высших растений – склеренхима и колленхима.
Склеренхима, как правило, состоит из клеток вытянутой формы – волокнообразных. Их клеточные стенки утолщаются и лигнифицируются, то есть одревесневают. Живое содержимое клетки впоследствии отмирает. Таким образом, склеренхима – это мертвая ткань, механическую функцию в которой выполняют жесткие клеточные стенки. Склеренхима твердая жесткая ткань и в растении она выполняет армирующую функцию, располагаясь обычно тяжами или слоями. Однако иногда склеренхима может быть представлена в виде отдельных клеток с одревесневшими клеточными стенками, разбросанных в толще некой мягкой ткани (например, паренхимы). Такие клетки называются склереидами. По форме различают разные типы склереид: брахисклереиды, астросклереиды, остеосклереиды и волокнистые склереиды. Все склеренхимные элементы вместе составляют стереом – совокупность всех толстостенных одревесневших клеток растения. Следует также помнить, что отчасти механическую функцию, подобно склеренхиме, выполняет водопроводящая ткань ксилема (в особенности ядровая древесина – вторичная ксилема, прекратившая проводить воду).
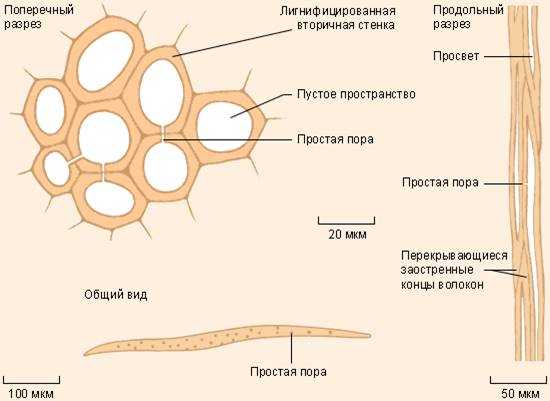
Рисунок 1: Склеренхима.
Колленхима также является механической тканью, однако клетки ее остаются живыми. Их клеточные стенки утолщаются, но неравномерно и не одревесневают. Живые клетки упругие, так как находятся под тургорным давлением, а клеточные стенки эластичны, поскольку состоят из полисахаридов. Именно эти свойства и позволяют колленхиме выполнять свою механическую функцию. Таким образом, колленхима – это живая упругая эластичная механическая ткань. Обычно колленхима располагается в тех органах высших растений, которые подвержены изгибу и должны быть упругими. Например, это стебли травянистых растений, особенно если стебель граненый или ребристый, то вдоль граней под эпидермой, скорее всего, располагаются тяжи колленхимы. Также колленхима часто встречается в листьях в черешке и вдоль средней жилки, поскольку именно эти части должны быть эластичными и упругими. Выделяют три типа колленхимы: уголковую (клеточные стенки утолщены в местах контакта трех и более клеток – «в уголках»), пластинчатую (утолщены продольные клеточные стенки) и рыхлую (похожа на уголковую, но с крупными межклетниками).
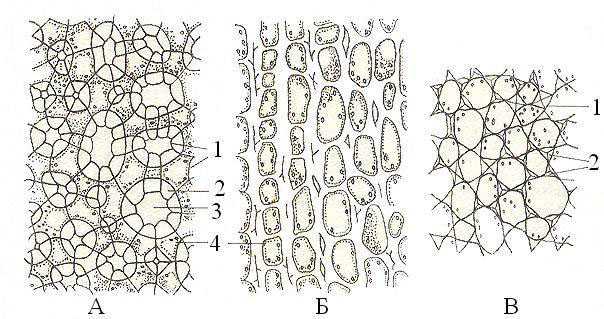
Рисунок 2: Колленхима. А – рыхлая; Б – пластинчатая; В – уголковая. 1 – первичная; клеточная стенка; 2 – вторичная клеточная стенка; 3 – межклетник; 4 – протопласт.
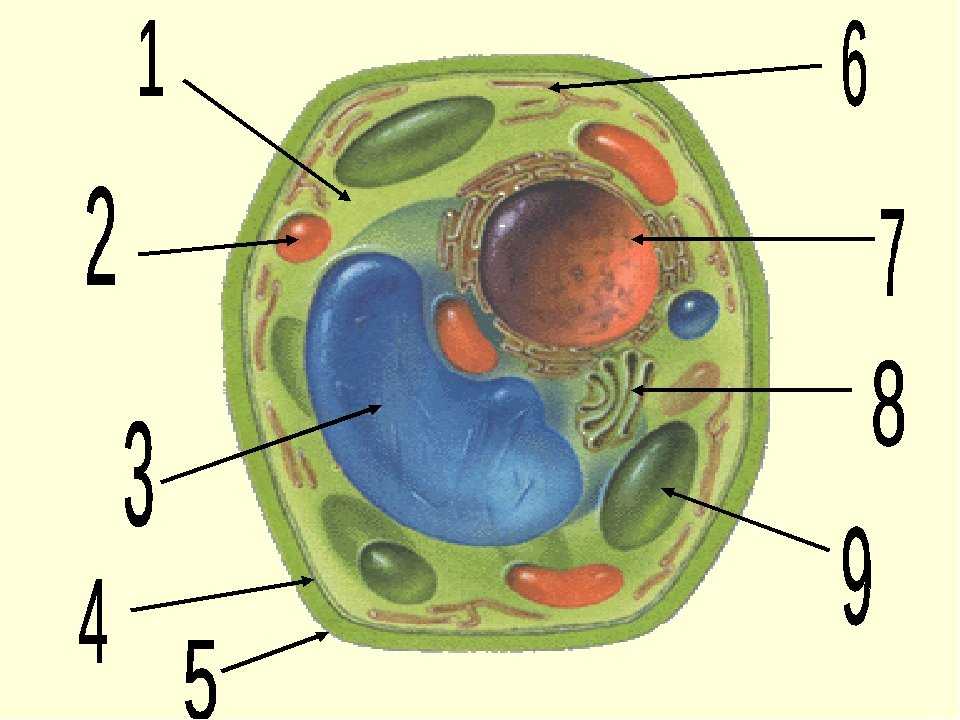
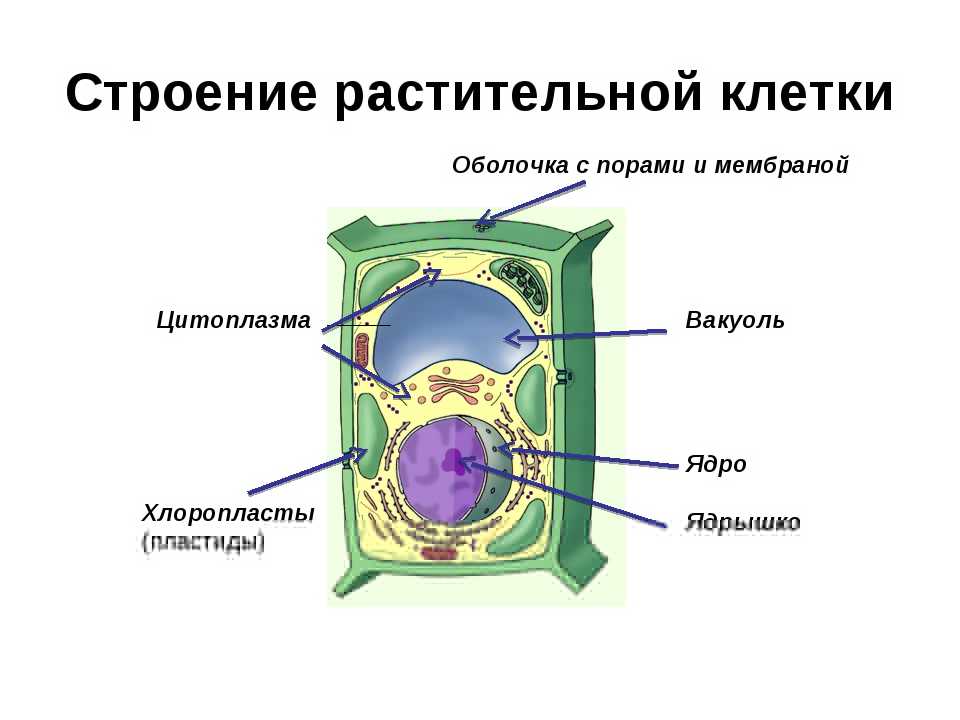
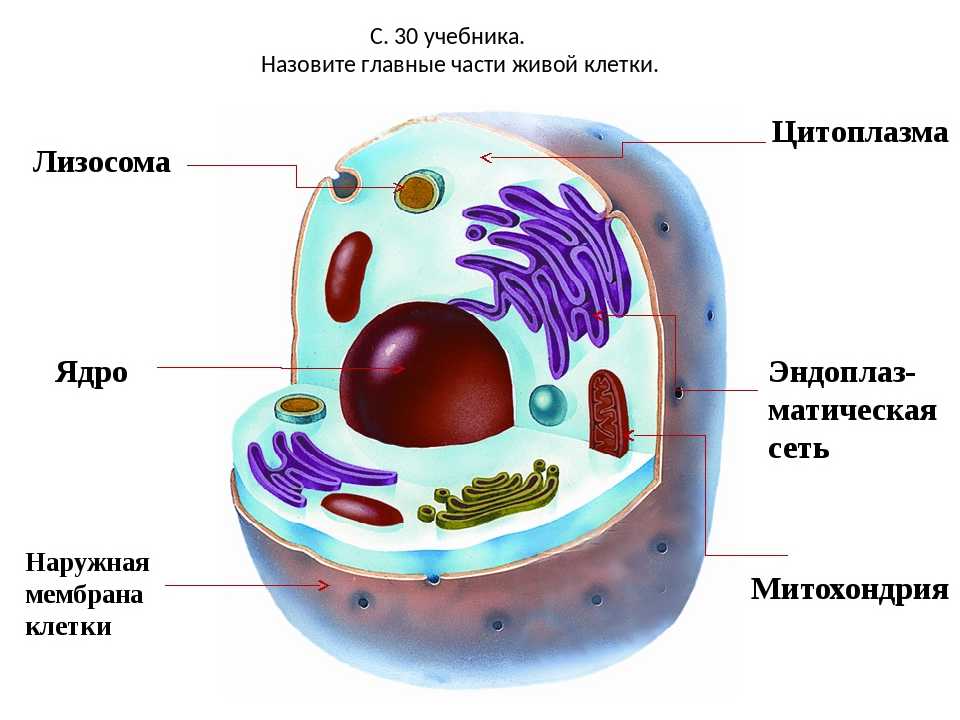
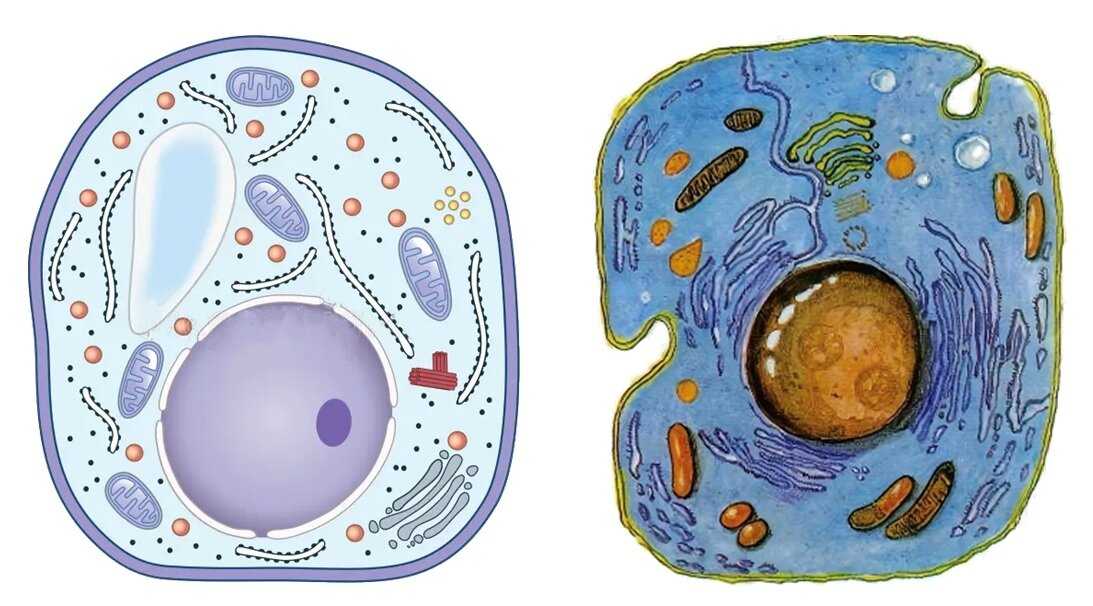
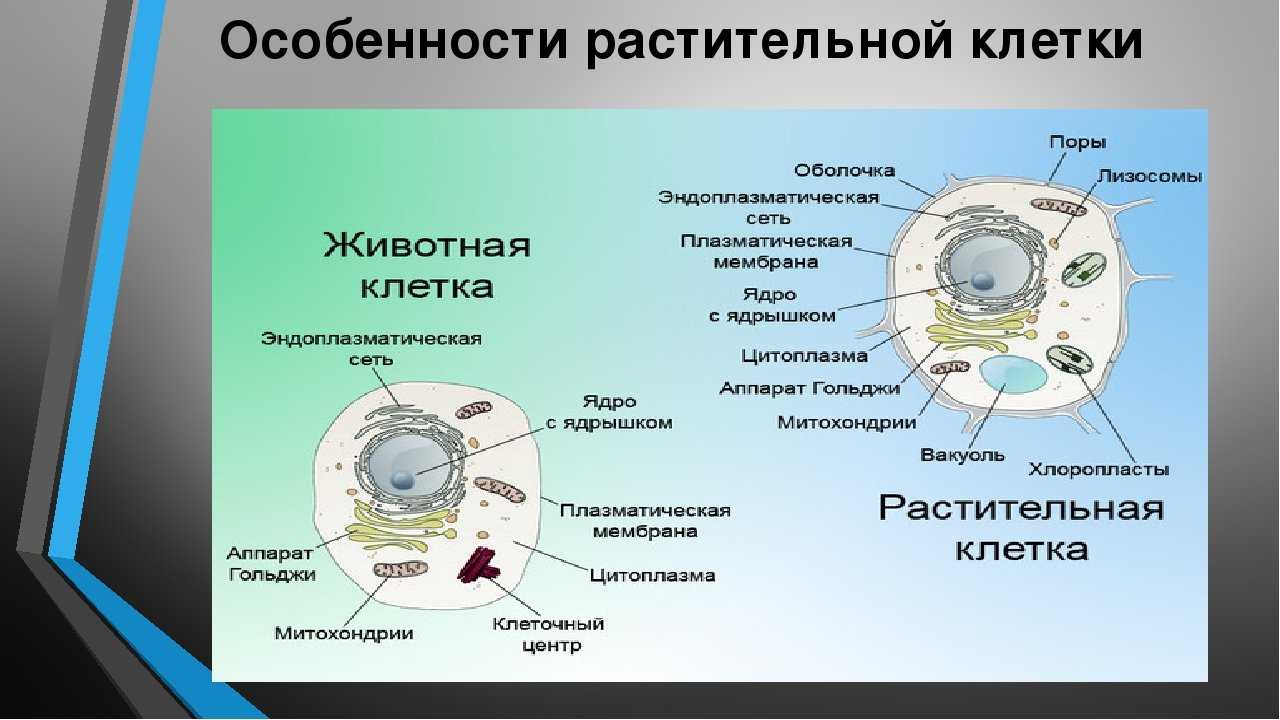
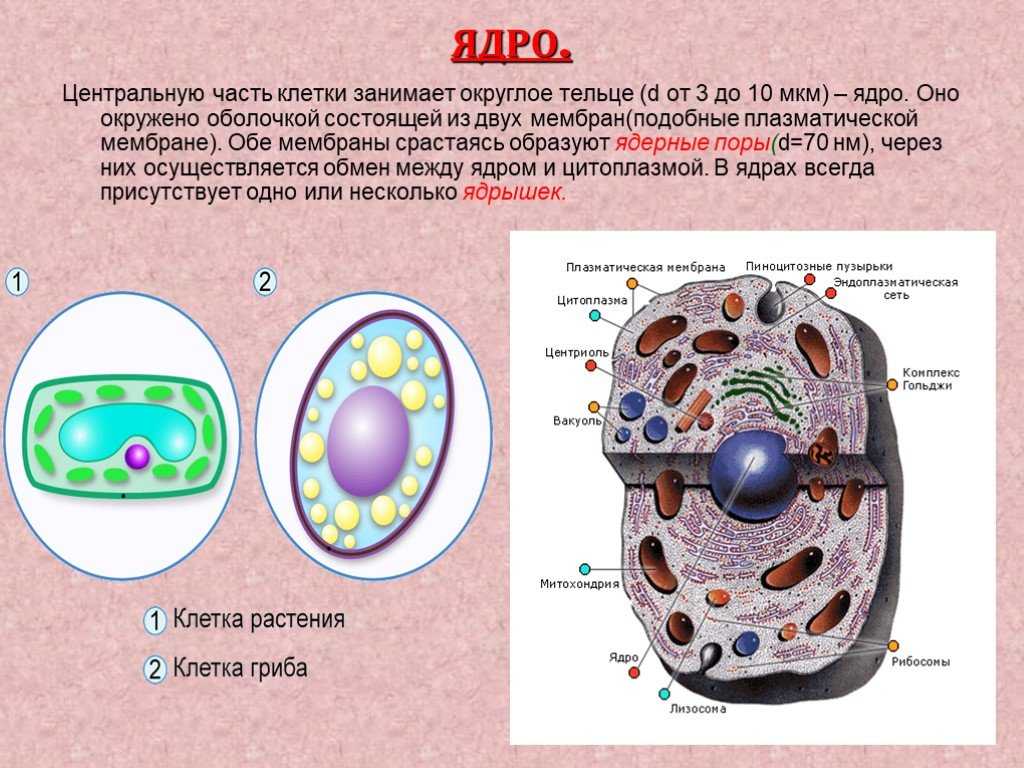
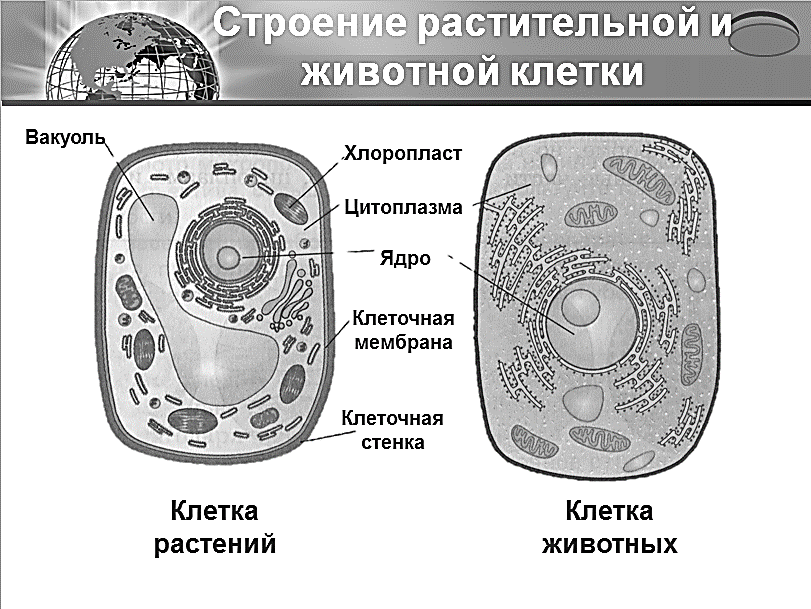
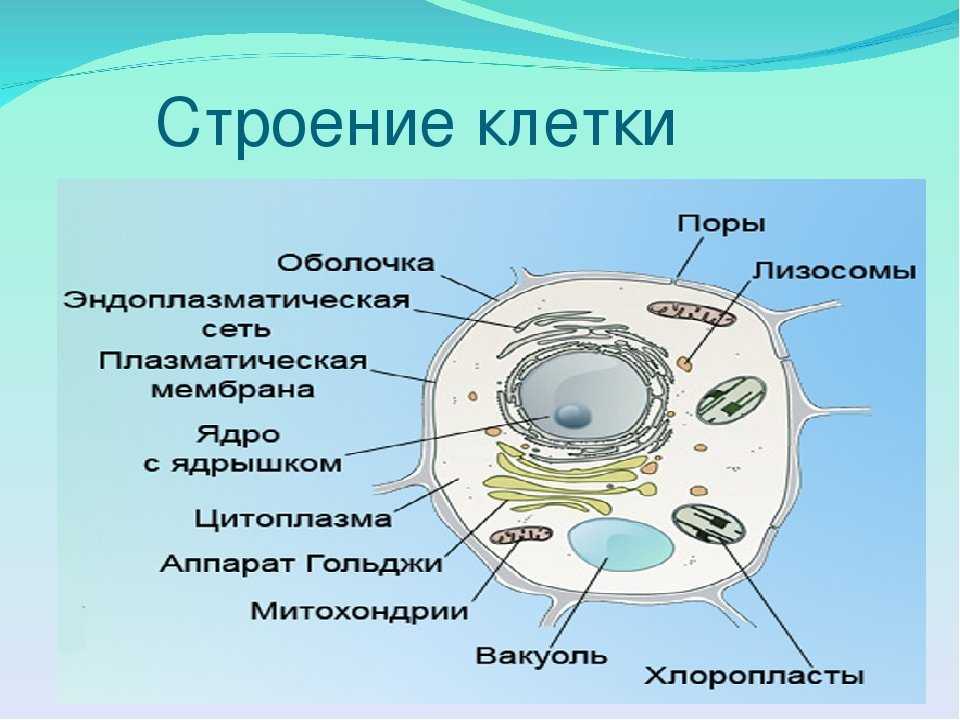
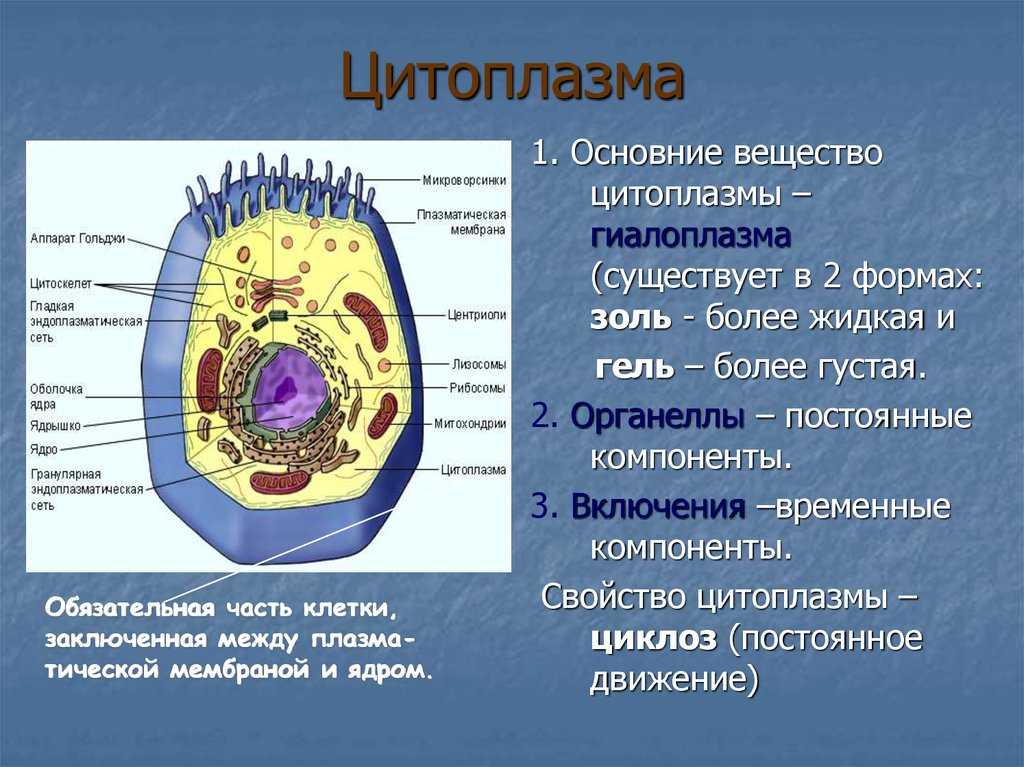
Клеточная мембрана: структура и функции
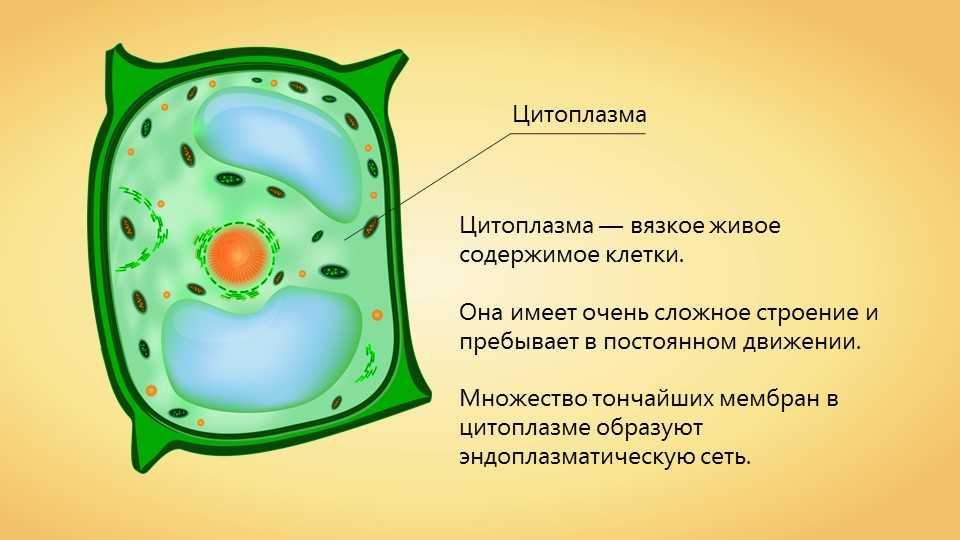
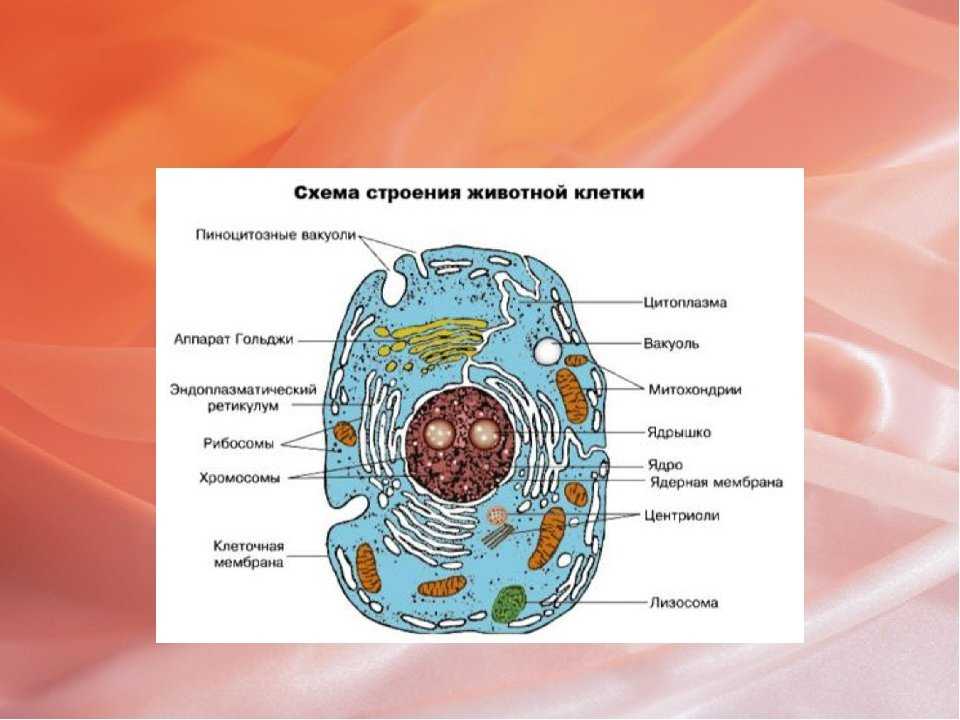
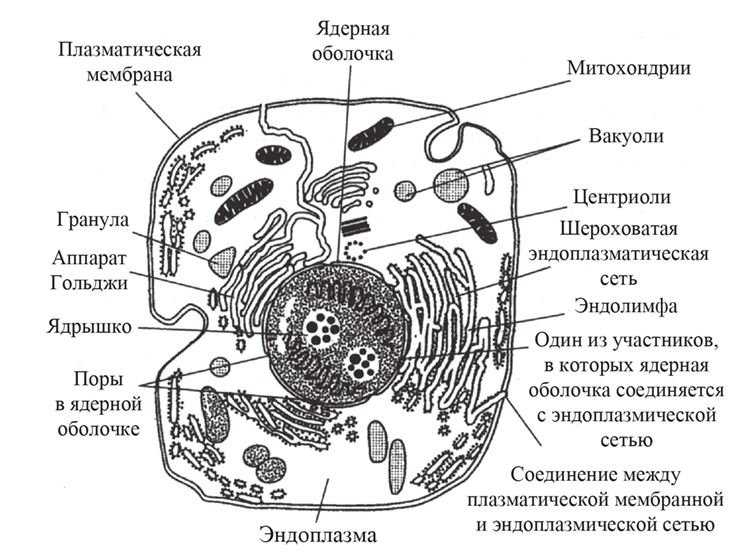
Клеточная мембрана — это тонкий слой, который образует границу между внутренним содержимым клетки и окружающей средой. Она состоит из липидного бислоя, включающего два слоя фосфолипидов и белковые каналы и рецепторы. Она служит для защиты клетки от вредных веществ, контролирует поступление и выведение веществ, а также участвует в межклеточном взаимодействии.
Фосфолипиды, находящиеся в мембране, упорядочены в двуслой. Внутренние «головки» фосфолипидов обращены друг к другу, а внешние лицевые поверхности связаны с водой. Кроме того, на мембране находятся белки, которые могут помочь фосфолипидам в их функциях и предоставить сообщения между ячейками.
Функции мембраны включают создание и поддержание градиента электролитов, которые нужны для жизни клетки, а также пропускание различных молекул через мембрану, чтобы способствовать обмену веществ между клеткой и внешней средой. Кроме того, мембрана может действовать как барьер между вредными веществами и клеткой, что помогает защитить клетку от ущерба.
В целом, мембрана играет важную роль в жизнедеятельности клетки и обеспечивает ее многомерный функционал.
Вопрос-ответ:
Что представляет собой клеточная мембрана?
Клеточная мембрана — это биологическая мембрана, которая разделяет внутреннюю среду клетки и внешнюю среду. Она состоит из двух слоев фосфолипидов, в которые встроены белки и гликолипиды.
Какова структура клеточной мембраны?
Клеточная мембрана состоит из двух слоев фосфолипидов, гидрофильные «головки» которых обращены наружу, а гидрофобные «хвосты» направлены друг к другу. В мембрану встроены белки, гликолипиды и холестерин.
Что такое перенос веществ через клеточную мембрану?
Перенос веществ через клеточную мембрану — это процесс, при котором различные вещества проходят через мембрану, чтобы попадать внутрь или наружу клетки. Существуют два типа переноса: пассивный (без затрат энергии) и активный (с затратой энергии).
Какие белки могут быть встроены в клеточную мембрану?
В клеточную мембрану могут быть встроены различные типы белков: рецепторы, каналы, транспортеры, ферменты, адаптеры, структурные белки и т.д. Каждый тип белков выполняет свою определенную функцию.
Как клетки могут взаимодействовать друг с другом через клеточную мембрану?
Клетки могут взаимодействовать друг с другом через клеточную мембрану с помощью специальных белков, которые находятся на поверхности мембраны. Они могут связываться друг с другом и передавать сигналы, что позволяет клеткам координировать свою деятельность.
Что такое гликолипиды и какова их роль в клеточной мембране?
Гликолипиды — это липиды, к которым присоединены углеводы. Они находятся на поверхности внешнего слоя клеточной мембраны и выполняют различные функции, такие как защита клетки от вредных веществ и участие в распознавании клеток других организмов.
Трансмембранный потенциал
Трансмембранный потенциал – это разность зарядов между внешней и внутренней сторонами клеточной мембраны, которая обеспечивает основные функции мембраны, такие как транспорт веществ, обмен веществ, передача сигналов.
Клеточная мембрана является барьером, который регулирует движение веществ, и трансмембранный потенциал — важный фактор, определяющий направление и число веществ, проходящих через мембрану. При снижении трансмембранного потенциала, более положительные ионы, такие как Na+, K+, перемещаются внутрь клетки, а отрицательные ионы – наружу.


В итоге, трансмембранный потенциал является необходимой составляющей клеточной мембраны, обеспечивающей ее основные функции. Его изменение может приводить к серьезным нарушениям жизнедеятельности клетки и приводить к различным патологиям.
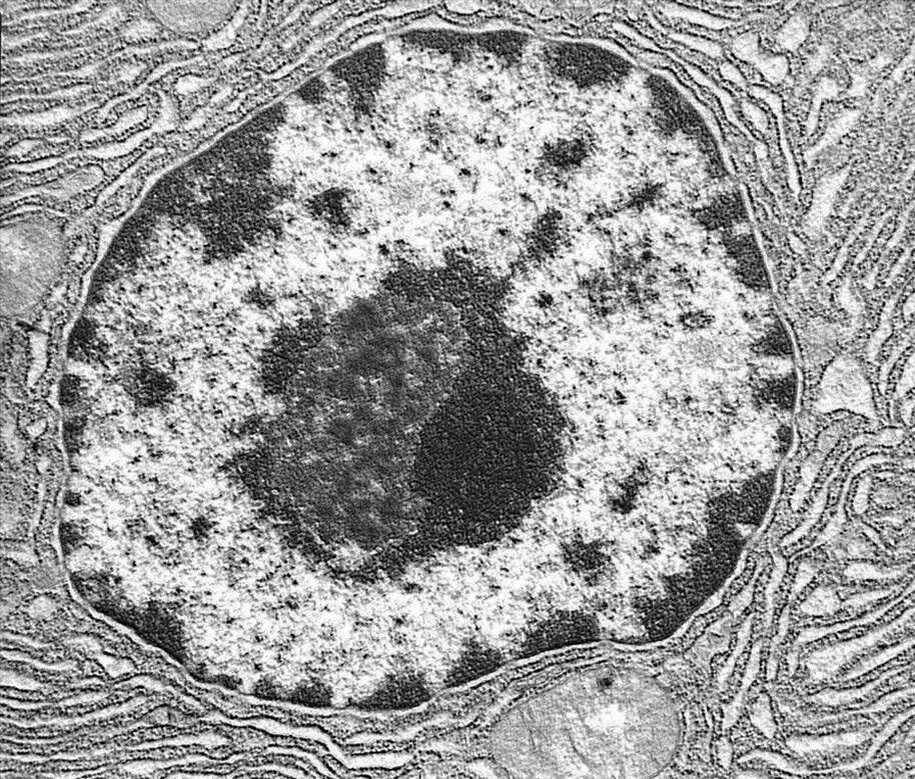
Высокопроницаемые (щелевые) контакты[править]
Щелевое соединение
Щелевое соединение, щелевой контакт — способ соединения клеток в организме с помощью белковых каналов (коннексонов). Через щелевые контакты могут непосредственно передаваться от клетки к клетке электрические сигналы (потенциалы действия), а также малые молекулы (с молекулярной массой примерно до 1.000 Д). Этим щелевые контакты отличаются от плазмодесм, через которые могут транспортироваться макромолекулы и даже органоиды.
Структурную основу щелевого соединения составляют коннексоны — каналы, образуемые шестью белками-коннексинами. В нервной системе щелевое соединение между нейронами встречается в так называемых электрических синапсах. Отдельные коннексоны обычно сосредоточены на ограниченных по площади участках мембран — нексусах, или бляшках (англ. plaque) диаметром 0,5-1 мкм. В области нексуса мембраны соседних клеток сближены, расстояние между ними составляет 2-4 нм.
Белки щелевых контактов
У позвоночных основу щелевых контактов составляют коннексины — первое из описанных семейств белков щелевых контактов. В геноме человека идентифицирован 21 ген щелевых контактов, в геноме мыши — 20 генов.
У беспозвоночных имеется другое семейство белков щелевых контактов, сходных с коннексинами по структуре и функциям. но негомологичных им (имеющих несходную первичную структуру) — иннексины. В геноме Caenorhabditis elegans найдено 25 генов иннексинов, в геноме Drosophila melanogaster — 8.
Позднее выяснилось, что у позвоночных, кроме коннексинов, имеются также белки, гомологичные иннексинам. Эти белки, открытые группой российских ученых под руководством Ю. В. Панчина, получили название паннексины. В геноме человека и мыши к настоящему времени идентифицированы 3 гена паннексинов.
У кишечнополостных и иглокожих есть щелевые контакты, но нет генов ни одного из вышеназванных семейств. Это означает, что существуют ещё не открытые семейства белков щелевых контактов.