

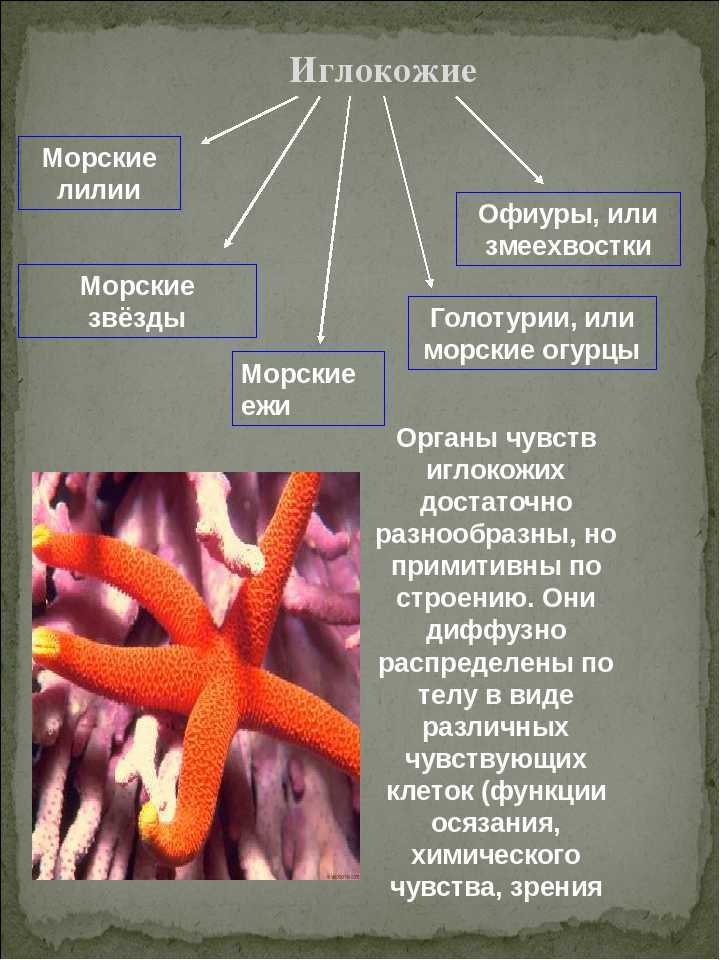
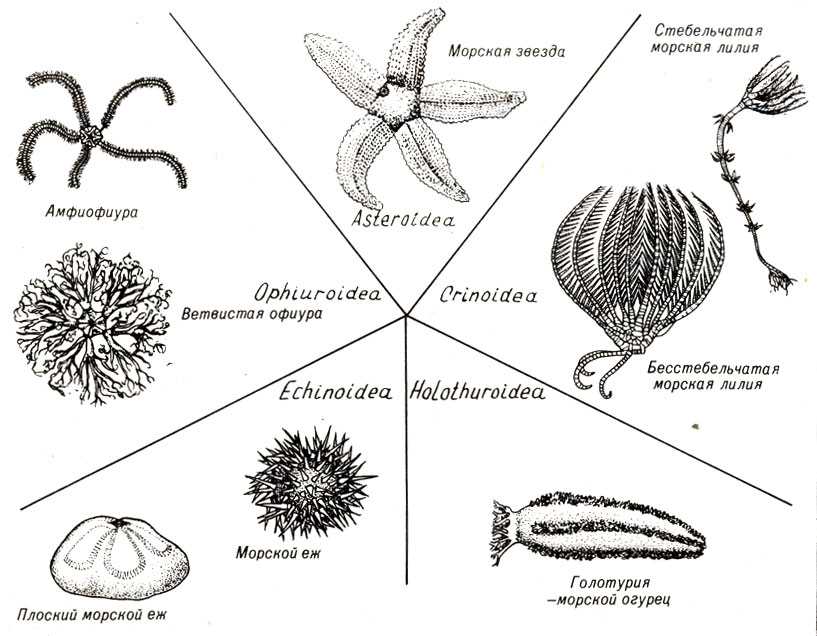
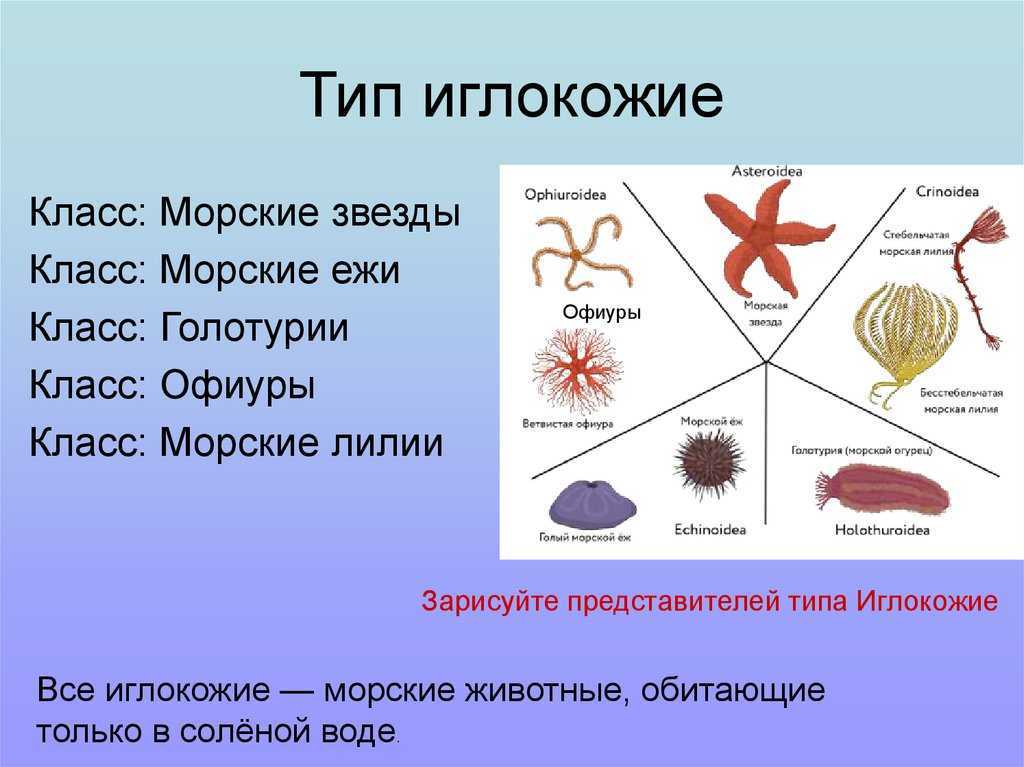
Классификация
Amplexograptus , полухордовый граптолит, обнаруженный в ордовике близ Кейни-Спрингс , штат Теннесси .
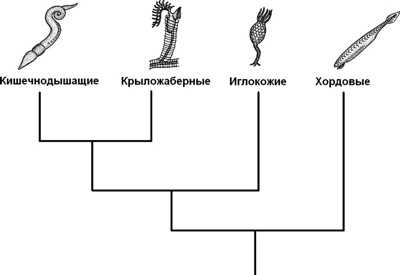
Hemichordata делятся на два класса: Enteropneusta , обычно называемые желудевыми червями, и Pterobranchia , которые могут включать граптолиты . Третий класс, Planctosphaeroidea , предложен на основе одного вида, известного только по личинкам. Тип насчитывает около 120 живых видов. Hemichordata, по-видимому, является сестрой иглокожих как Ambulacraria; Xenoturbellida может быть базальной по отношению к этой группе. Птеробранхии могут происходить из Enteropneusta, что делает Enteropneusta парафилетическим. Возможно, что вымерший организм Etacystis является членом Hemichordata либо внутри, либо с близким родством с Pterobranchia.
Существует 130 описанных видов Hemichordata, и обнаруживается много новых видов, особенно в морских глубинах.
Филогения
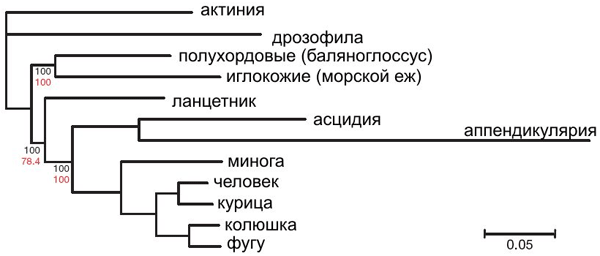
Филогенетическое дерево , показывающее положение полухордовых является:
| Deuterostomia |
|
||||||||||||||||||||||||
Внутренние отношения внутри полухордовых показаны ниже. Дерево основано на данных о последовательности 16S + 18S рРНК и филогеномных исследованиях из множества источников.
| Hemichordata |
|
||||||||||||||||||||||||||||||||||||||||||||||||
Охрана
16 видов иглокожих занесены в Красный список угрожаемых видов МСОП (The IUCN Red List of Threatened Species) как редкие и исчезающие, из них 9 видов — уязвимые и 7 — вымирающие. Все 16 являются голотуриями (морскими огурцами) из семейств Holothuriidae и Stichopodidae отряда Aspidochirotida.
Actinopyga echinites Actinopyga mauritiana Actinopyga miliaris Apostichopus japonicus — Дальневосточный трепанг Apostichopus parvimensis Bohadschia maculisparsa Holothuria arenacava Holothuria fuscogilva Holothuria lessoni Holothuria nobilis Holothuria platei Holothuria scabra Holothuria whitmaei Isostichopus fuscus Stichopus herrmanni Thelenota ananas
Происхождение и эволюция
Благодаря минерализированному скелету иглокожие хорошо сохраняются в ископаемом состоянии (хотя и часто представлены лишь разрозненными скелетными элементами), а своеобразная структура этих элементов (стереом) даёт возможность легко их опознать. Древнейшие известные (на 2014 год) ископаемые остатки иглокожих относятся к третьему веку кембрия (около 515—520 млн лет назад), когда они примерно одновременно появляются в морях Лаврентии, Ангариды и Гондваны. Это представители геликоплакоидей, эдриоастероидей и эокриноидей. Гипотезы о принадлежности к иглокожим некоторых докембрийских организмов (например, Arkarua) не получили широкой поддержки. Представители пяти современных классов известны начиная с нижнего — среднего ордовика. Расцвет типа приходится на палеозой.
Ближайшие родственники иглокожих среди современных животных — полухордовые, образующие вместе с ними кладу Ambulacraria. Эта клада является сестринской группой хордовых. Время разделения полухордовых и иглокожих методом молекулярных часов оценивают примерно в 580—550 млн лет назад.
Общим предком всех вторичноротых было билатерально-симметричное свободноживущее животное с тремя парами целомических мешков. На это указывает наличие стадии развития, сходной у всех вторичноротых. У иглокожих данной стадии соответствует личинка диплеврула. Появление первых иглокожих связано с переходом этого гипотетического предка к сидячему образу жизни и приобретением им радиальной симметрии.
В пятом веке кембрия появились представители класса Carpoidea, просуществовавшего до нижнего девона. Вели сидячий образ жизни, но ещё не обладали радиальной симметрией. Тело было покрыто пластинками, рот и анус размещались на стороне, обращённой от субстрата. Внутренние органы располагались асимметрично. У представителей класса Cystoidea (шаровики) вокруг рта появились радиальные амбулакральные бороздки, предназначенные для собирания пищи из толщи воды. От шаровиков ведут начало остальные Pelmatozoa: класс Blastoidea (морские бутоны), современные морские лилии и класс Edrioasteroidea, в составе которого появились свободноживущие виды.
Первые Eleutherozoa, сочетающие особенности современных морских звёзд, офиур и морских ежей, относились к классу Ophiocistia. От них и произошли современные представители подтипа.
У всех иглокожих наблюдается инверсия первых трех HOX-генов и их транслокация в конец HOX-кластера по сравнению с остальными вторичноротыми. Передние HOX-гены принимают участие в развитии головы (у позвоночных они экспрессируются в продолговатом мозге). У иглокожих нет ни головы, ни головного мозга. Характер причинно-следственной связи между инверсией/транслокацией этих генов у иглокожих и их переходом к лучевой симметрии в настоящее время является дискуссионным.





Кладограмма современных классов иглокожих по молекулярным данным согласно A. Reich с соавт., 2015:
Ссылки
- Аламо, М.А.Ф., и Ривас, Г. (ред.). (2007). Уровни организации у животных. UNAM.
- Кэмерон, К. Б. (2005). Филогения полухордовых по морфологическим признакам. Канадский журнал зоологии, 83(1), 196-215.
- Хикман, К. П., Робертс, Л. С., Ларсон, А., Обер, В. К., и Гаррисон, К. (2001). Интегральные принципы зоологии. Макгроу-Хилл.
- Ирвин, М. Д., Стоунер, Дж. Б., и Кобау, А. М. (ред.). (2013). Зоопарк: введение в науку и технологии. Издательство Чикагского университета.
- Маршалл, А. Дж., И Уильямс, В. Д. (1985). Зоология. Беспозвоночные (Том 1). Я поменял направление.
- Паркер, Т. Дж., И Хасуэлл, В. А. (1987). Зоология. Хордовые (Том 2). Я поменял направление.
- Сато Н., Тагава К., Лоу, К. Дж., Ю, Дж. К., Кавашима, Т., Такахаши, Х.,… и Герхарт, Дж. (2014). О возможной эволюционной связи стомохорды полухордовых с глоточными органами хордовых. Бытие, 52(12), 925-934.
- Тассия, М. Г., Кэннон, Дж. Т., Коникофф, К. Э., Шенкар, Н., Халаныч, К. М., и Свалла, Б. Дж. (2016). Глобальное разнообразие Hemichordata. PloS один, 11(10), e0162564.
Кто относится к первичноротым и вторичноротым?
Животные делятся на первичноротых и вторичноротых по способу развития рта. Организмы, у которых первичный рот зародыша становится ртом взрослой формы называются первичноротыми.
К первичноротым относятся большинство беспозвоночных: кишечнополостные, плоские, круглые и кольчатые черви, моллюски и членистоногие.
В случае если рот взрослой формы возникает не из первичного рта зародыша, а прорывается на обратной стороне — такие животные называются вторичноротыми. Фактически у вторичноротых в процессе эмбриогенеза рот и анальное отверстие меняются местами.
Вторичноротыми являются хордовые, а также иглокожие (морские звезды, морские ежи).
P.S. Мы нашли статью, которая относится к данной теме, изучите ее — Животные
P.S.S. Для вас готов следующий случайный вопрос. Мы сами не знаем, но вас ждет что-то интересное!
Беллевич Юрий Сергеевич 2018-2021
Текст и опубликованные материалы являются интеллектуальной собственностью Беллевича Юрия Сергеевича. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов вопроса и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Источник
Класс Голотурии (Holothuroidea)
Голотурии (морские огурцы) обладают мешковидным или червеобразным телом. Скелет подвергается редукции и представлен отдельными известковыми пластинками (спикулами). Радиальная пятилучевая симметрия выражена слабо: большинство органов расположены билатерально. Животные ползают на брюшной стороне. Рот находится на переднем полюсе тела, окружен венчиком простых или ветвящихся щупалец. Щупальца представляют собой видоизмененные амбулакральные ножки. С помощью щупалец голотурии захватывают ил или ловят мелких животных. На заднем полюсе тела находится анус.
От переднего конца тела к заднему тянутся пять амбулакральных полосок. Три из них лежат на нижней стороне тела (тривиум), а две — на верхней стороне тела (бивиум). Голотурии — малоподвижные донные животные, обитают обычно зарывшись в ил. Есть виды, ведущие планктонный образ жизни. При раздражении могут сильно сжиматься, втягивая при этом внутрь переднюю часть тела со щупальцами.
Кишечник в виде трубки тянется вдоль тела, в задней части расширяется и образует мускулистую клоаку. В клоаку открываются особые органы, имеющие защитное значение. Это железистые трубочки, которые при раздражении животного выбрасываются наружу через отверстие клоаки и превращаются в белые липкие нити, облепляющие нападающего врага. В клоаку также открываются водяные легкие — органы дыхания голотурий. Легкие — это тонкостенные ветвящиеся мешки, вода в которые нагнетается через клоакальную щель, а затем выбрасывается тем же путем.
Выделение происходит с помощью амебоцитов. Половая система образована одной железой, проток которой открывается на поверхности тела. Голотурии могут быть как раздельнополыми, так и гермафродитами. Оплодотворение наружное, развитие протекает с метаморфозом.
При раздражении голотурии могут выбрасывать через разрыв стенки клоаки свои внутренности. Этот процесс называется эвисцерацией. Через некоторое время животные регенерируют утраченные органы. Некоторые голотурии способны к автотомии: они отшнуровывают заднюю часть туловища, а затем ее восстанавливают.
Промысловое значение имеют трепанги (Stichopus) и кукумарии (Cucumaria).
Эволюционное место и происхождение вторичноротых животных
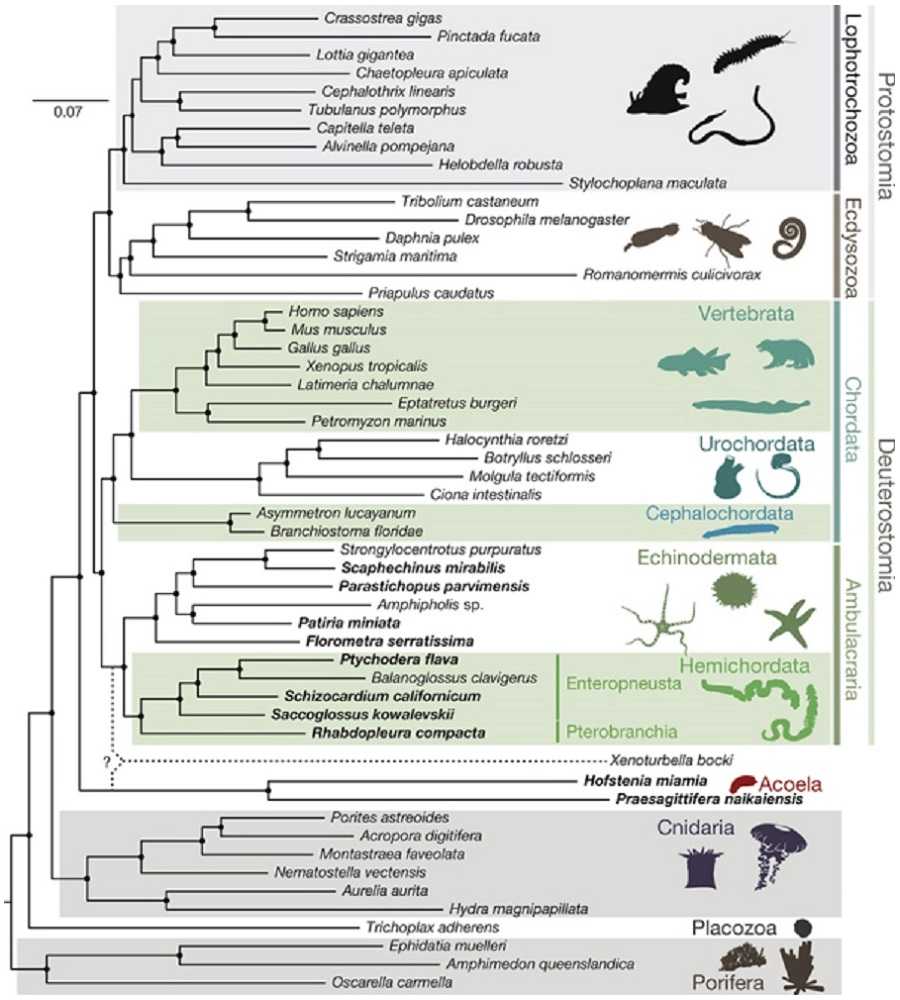
Филогенетическое дерево вторичноротых животных
 |
Раньше считалось, что в группу вторичноротых входят 3 типа животных:
Теперь в объединение вторичноротых включают также щетинкочелюстных и погонофор, |
Вторичноротые животные (дейтеростомы)
/ | |
/ / \ / \
/ / \ / \
Щетинкочелюстные Полухордовые Иглокожие Погонофоры Хордовые
|
|-перистожаберные (крыложаберные)
|
|-кишечнодышащие
|
|-граптолиты (ископаемые)
|
Гипотезы происхождения вторичноротых животных
|
Происхождение вторичноротых неясно.
У вторичноротых в период зародышевого развития на месте первичного рта (бластопора) образуется анальное отверстие,
Все билатерально-симметричные организмы делятся на две группы — первичноротые и вторичноротые. Так, у первичноротых животных бластопор (возникающее в эмбриональном развитии отверстие первичного кишечника) У вторичноротых (например, у иглокожих) бластопор становится анальным отверстием, Помимо судьбы рта, первичноротые и вторичноротые отличаются другими эмбриологическими особенностями. (1) Во-первых, это характер дробления оплодотворенного яйца: (2) Другое отличие состоит в способе закладке целома (вторичной полости тела): (3) Третьим важнейшим различием между первично- и вторичноротыми является судьба зачатка первичного мозга: |


![Тип echinodermata. иглокожие [1974 друщиц в.в. - палеонтология беспозвоночных]](https://lesniepolyani.ru/wp-content/uploads/b/8/a/b8a8bf0403308a53180c27076d022424.jpeg)







Высказана гипотеза о происхождении вторичноротых от первичноротых.
Образ жизни и некоторые черты строения предка вторичноротых
Общий предок вторичноротых животных, по-видимому, вел донный образ жизни,
ползая на брюшной стороне с помощью ресничной полоски, подостланной вентральным нервным тяжем.
Он имел три пары целомических мешков и щупальцевый аппарат, с помощью которого собирал мелкие пищевые частицы.
В дальнейшем могло произойти обособление самого заднего отдела тела,
с помощью которого предки вторичноротых временно зарывались в верхние слои грунта
В эволюции вторичноротых этому отделу принадлежит очень важное место.
Очевидно, у него уже были особые отверстия, соединяющие полость глотки с внешней средой: жаберные поры.
Он был фильтратором и питался бактериями и органической взвесью, которая попадала в глотку с водой через рот
и осаждалась на слизистом эпителии, а вода выходила наружу через жаберные поры.
Именно так и сегодня живут примитивные вторичноротые (кишечнодышащие, ланцетник, личинки миног),
и такой же способ питания предполагается для древнейших ископаемых вторичноротых, таких как юннанозои и пикайя.
Разнообразие хордовых

Данный тип включает в себя три подтипа: Бесчерепные, Личиночнохордовые (Оболочники) и Черепные (Позвоночные). К первому из них относится только 30 видов, встречающихся в наше время. Их представителями являются ланцетники. Эти животные внешне напоминают хирургический инструмент под названием ланцет. Тело этих небольших животных практически всегда наполовину находится в песке.
Так ланцетнику удобнее фильтровать воду, заглатывая питательные частицы. Самым многочисленным подтипом хордовых являются Позвоночные. Они освоили абсолютно все среды обитания, заполнили пищевые цепи и экологические ниши. Водными обитателями являются рыбы. Их тело обтекаемой формы покрыто чешуей, они приспособлены к жаберному дыханию, двигаются при помощи плавников.
Первыми вышедшими на сушу являются земноводные животные. Это лягушки, жабы, тритоны, черви и рыбозмеи. Их общее название связано с тем, что они живут на суше, дышат при помощи легких и кожи, а вот процесс их размножения происходит в воде. Подобно рыбам, их самки мечут в воду икру, которую самцы поливают семенной жидкостью.

Типично наземные животные – это пресмыкающиеся. Ящерицы, змеи, черепахи и крокодилы проводят в воде лишь время охоты. Размножаются они яйцами, которые откладывают в особых укрытиях на суше. Их кожа сухая, покрыта плотными чешуйками. Последний признак унаследовали от пресмыкающихся птицы. Неоперенная часть их ног называется цевка. Именно она и покрыта небольшими чешуйками. Этот факт ученые рассматривают как доказательство происхождения в процессе эволюции.
Птицы способны к полету благодаря многим чертам внешнего и внутреннего строения. Это видоизмененные передние конечности, перьевой покров, легкий скелет, наличие киля – плоской кости, к которой крепятся мышцы, приводящие в движение крылья.
Наконец, Звери, или Млекопитающие, являются венцом эволюции. Они живородящие и выкармливают детенышей молоком. Хордовые животные – самые сложноорганизованные, разнообразные по строению, играющие важнейшую роль в природе и жизни человека.
Морфология
Изначально вторичноротые были объединены на основании данных сравнительной эмбриологии, поскольку ранние этапы эмбриогенеза многих хордовых, полухордовых и иглокожих демонстрируют общие черты:
- Радиальное дробление. Когда зигота начинает делиться, первые две плоскости дробления проходят через анимальный и вегетальный полюса зиготы (то есть по «меридианам»), перпендикулярно друг другу. Третья проходит через экватор. При дальнейших делениях эмбрион сохраняет сферическую симметрию. В конце концов он превращается в бластулу — полый шарик, заполненный жидкостью
- Гаструляция путём инвагинации. Клетки, находящиеся на вегетальном полюсе бластулы, вворачиваются внутрь, давая начало эндомезодерме. Они формируют первичную кишку — архентерон, который открывается наружу отверстием, называемым бластопором.
- Судьба бластопора («первичного рта»). У большинства (хотя известно множество исключений) первичноротых билатерий бластопор превращается в ротовое отверстие либо закрывается. У вторичноротых бластопор чаще всего превращается в анус. Рот образуется на противоположном (анимальном) полушарии эмбриона, где архентерон приходит в соприкосновение с эктодермой.
- Источник мезодермы. Мезодерма образуется из выпячиваний архентерона, которые отшнуровываются и превращаются в целомические мешки. Данный процесс называется энтероцелией, в противовес шизоцелии, когда целомические полости образуются из неупорядоченного скопления клеток, находящихся между экто- и эндодермой. У иглокожих и полухордовых образуется три пары целомических мешков, расположенных симметрично друг относительно друга. Они носят названия прото-, мезо- и метацеля. У примитивных хордовых (например, ланцетников) пар целомических мешков (сомитов) образуется гораздо больше. Передние несколько пар образуются одновременно путём энтероцелии, задние — один за другим появляются в хвостовом конце тела животного путём шизоцелии.
У всех вторичноротых существует циркуляторная система, которая в наиболее примитивной форме состоит из спинного и брюшного сосуда, перекачивающих гемолимфу в задне-переднем и передне-заднем направлении, соответственно. У хордовых спинная и брюшная стороны тела, скорее всего, поменялись местами, и уже у ланцетников роль перистальтического насоса играет брюшной сосуд, качающий гемолимфу вперед. У общего предка позвоночных и оболочников впервые появляется полноценное сердце, которое, скорее всего, негомологично сердцу полухордовых.
Вероятные ископаемые предки вторичноротых организмов
«Лиловый носок» (Xenoturbella) — самое примитивное вторичноротое животное

Ксенотурбелла (лаь. Xenoturbella) — род морских червеобразных животных до 4 см в длину, выделяемый в отдельный тип Xenoturbellida. Известны также по имени «лиловый носок», потому что похожи именно на брошенный на пол носок.
Существа эти настолько необычны, что в течение 60 лет ученые не могли определить, к какому именно виду или роду они принадлежат.
Сквозной пищеварительный тракт отсутствует — имеется только мешкообразная кишка без заднего прохода, У них также нет централизованного мозга или нервного узла. Половой системы тоже нет, но обнаружены гаметы — репродуктивные клетки, имеющие гаплоидный набор хромосом. |
«Сморщенный мешок с короной» (Saccorhytus coronarius) — древнейшее вторичноротое?

Исследуя породы необычных сланцев в провинции Шаньси, которые сформировались во время «кембрийского взрыва»
Эти организмы, как предполагают ученые, обладали неким подобием жабр, следы которых можно заметить |
Древнейший предок людей оказался похожим на зубастый «мешок».
У Saccorhytus coronarius нет вообще анального отверстия. Рот усеян зубами.
Подтипы
Головохордовые
Головохордовое Branchiostoma lanceolatum Основная статья: Головохордовые
Головохо́рдовые
(лат. Cephalochordata) илибесчерепны́е (лат. Acrania) — небольшие морские рыбообразные животные со всеми признаками, свойственными хордовым. Бесчерепные — подтип низших хордовых животных, в отличие от других хордовых (оболочников и позвоночных), сохраняющий основные признаки типа (хорда, нервная трубка и жаберные щели) в течение всей жизни. Головной мозг отсутствует, органы чувств примитивны. Ведут придонный образ жизни, по характеру питания — роющие фильтраторы. Возможно являются предками позвоночных, либо являются последними живыми членами группы, от которой произошли позвоночные. Всего к бесчерепным относятся около 30 видов, составляющих один класс — ланцетники.









Оболочники
Оболочники: асцидии Основная статья: Оболочники
Оболо́чники
(лат. Tunicata, Urochordata) — подтип хордовых животных. Включают 5 классов — асцидий, аппендикулярий, сальп, огнетелок и бочёночников. По другой классификации последние 3 класса считаются отрядами класса Thaliacea. Известно более 1 000 видов. Большинство из них имеют хорду только на личиночной стадии. Они распространены по всему миру и населяют морское дно.
Три крупных класса оболочников:
- Асцидии — низшие мягкотелые хордовые-фильтраторы, во взрослом состоянии ведущие сидячий образ жизни;
- аппендикулярии сохраняют личиночные черты, такие как хвост, на протяжении всей жизни. По этой причине долгое время рассматривались как личинки асцидий и сальп. Из-за наличия длинных хвостов личинки оболочников называют лат. urochordata;
- третья группа оболочников — свободноплавающие сальпы, питаются планктоном. В их жизненном цикле известно два поколения — одиночное гермафродитное и почкующееся колониальное бесполое. У личинок этих животных есть все основные признаки хордовых, в том числе хорда и хвост. Они также оснащены рудиментарным мозгом и датчиками освещённости и положения (крена).
Позвоночные
Прыжки гребнистых крокодилов — город Дарвин, Австралия Основная статья: Позвоночные
Позвоно́чные
(лат. Vertebrata) — высший подтип хордовых животных. Доминирующая (наряду с насекомыми) на земле и в воздушной среде группа животных. Отличаются от других хордовых наличием обособленного черепа и развитием головного мозга и органов чувств. Хорда у большинства представителей высших хордовых замещается на позвоночник, защищающий спинной мозг и состоящий, как правило, из хрящевой и костной ткани. Эндостиль, как таковой, присутствует только у личинок миног. По сравнению с низшими хордовыми — бесчерепными и оболочниками — они характеризуются значительно более высоким уровнем организации, что наглядно выражено как в их строении, так и в физиологических отправлениях. Среди позвоночных нет видов, ведущих сидячий (прикреплённый) образ жизни. Они перемещаются в широких пределах, активно разыскивая и захватывая пищу, находя для размножения особей другого пола, спасаясь от преследования врагов.
Положение миног неоднозначно. У них в наличии недоразвитый череп и рудиментарные позвонки — следовательно, они могут рассматриваться как позвоночные и истинные рыбы. Тем не менее, молекулярные филогенетики, использовавшие биохимические реакции для классификации организмов, в результате эту группу позвоночных объявили сестринской семейству миксиновых
(лат. Myxinidae) класса круглоротых. Миксины, имеющие жаберный скелет, состоящий из небольшого числа хрящевых пластинок, рудиментарные позвонки, не рассматриваются как истинные позвоночные — их считают группой, из которой эволюционировали позвоночные.
Особенности строения
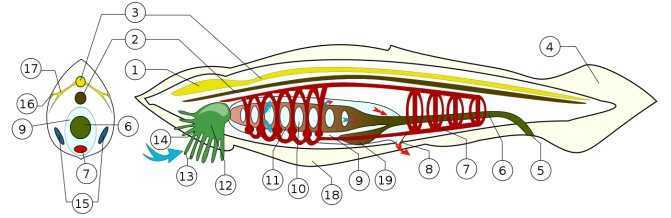
Схема строения головохордовых на примере Ланцетника: 1 — утолщение нервной трубки спереди («мозг»); 2 — хорда; 3 — спинной нервный тяж («спинной мозг»); 4 — хвостовой плавник; 5 — анус; 6 — пищеварительный канал; 7 — кровеносная система; 8 — выходное отверстие околожаберной полости (атриопор); 9 — околожаберная полость; 10 — глоточные (жаберные) щели; 11 — глотка; 12 — ротовая полость; 13 — околоротовые щупальца; 14 — ротовое отверстие; 15 — половые железы (семенники или яичники); 16 — глазки Гессе; 17 — нервы; 18 — метаплевральная складка; 19 — слепой печёночный вырост. Дыхание (газообмен): синей стрелкой указан вход воды, богатой кислородом, а красными — выход обогащённой углекислым газом. Хордовые — тип животных, характеризующихся билатеральной симметрией и наличием, по крайней мере, на определенных стадиях развития следующих признаков:
- Хорда, представляющая собой эластичный стержень энтодермального происхождения, образующийся путём выпячивания спинной стороны кишечной трубки. У позвоночных хорда в ходе эмбрионального развития полностью или частично замещается хрящевой и костной тканью, образующей позвоночник.
- Нервная трубка, расположенная дорсально. У позвоночных развивается в спинной мозг и головной мозг.
- Жаберные щели — парные отверстия в глотке. У низших хордовых участвуют в фильтрации воды для питания. У наземных позвоночных жаберные щели закладываются в раннем эмбриогенезе в виде жаберных мешочков.
- Мышечный хвост — постанальный отдел тела, расположенный каудальнее смещенного на брюшную сторону тела ануса (в него заходят хорда и нервная трубка, но не заходит кишечник).
- Эндостиль — желобок на вентральной стороне глотки. У низших хордовых-фильтраторов в нём производится слизь, помогающая собирать частицы пищи и доставлять их в пищевод. Также в нём накапливается иод и, возможно, он является предшественником щитовидной железы позвоночных. Как таковой, эндостиль у позвоночных есть только у пескоройки.