
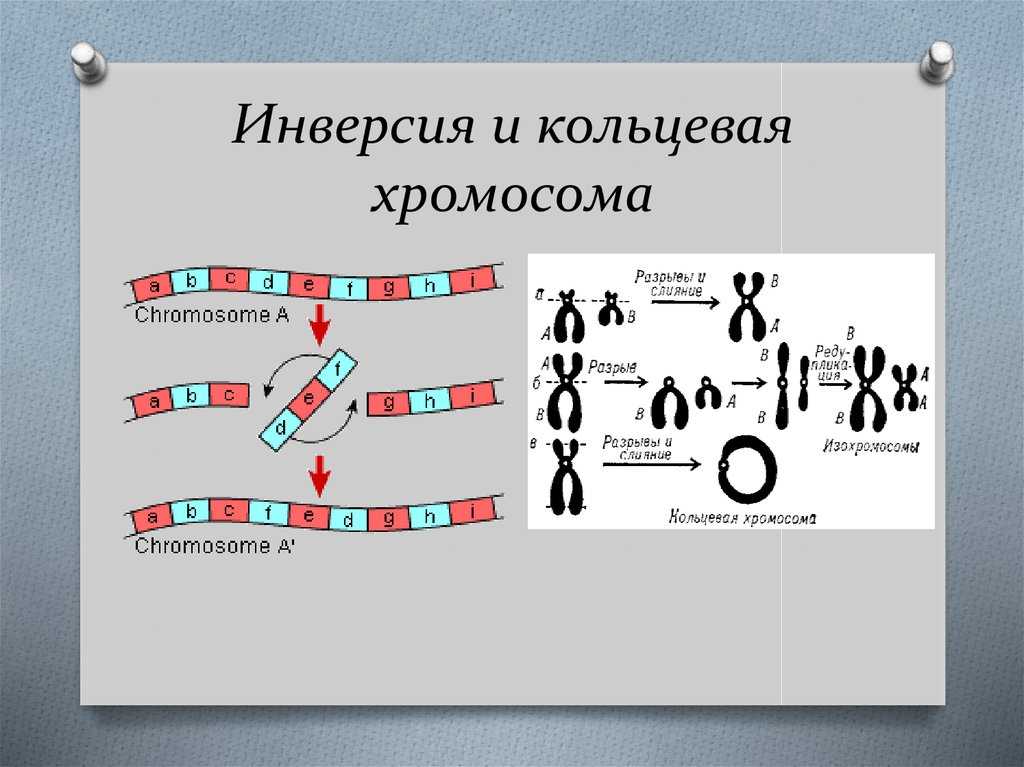
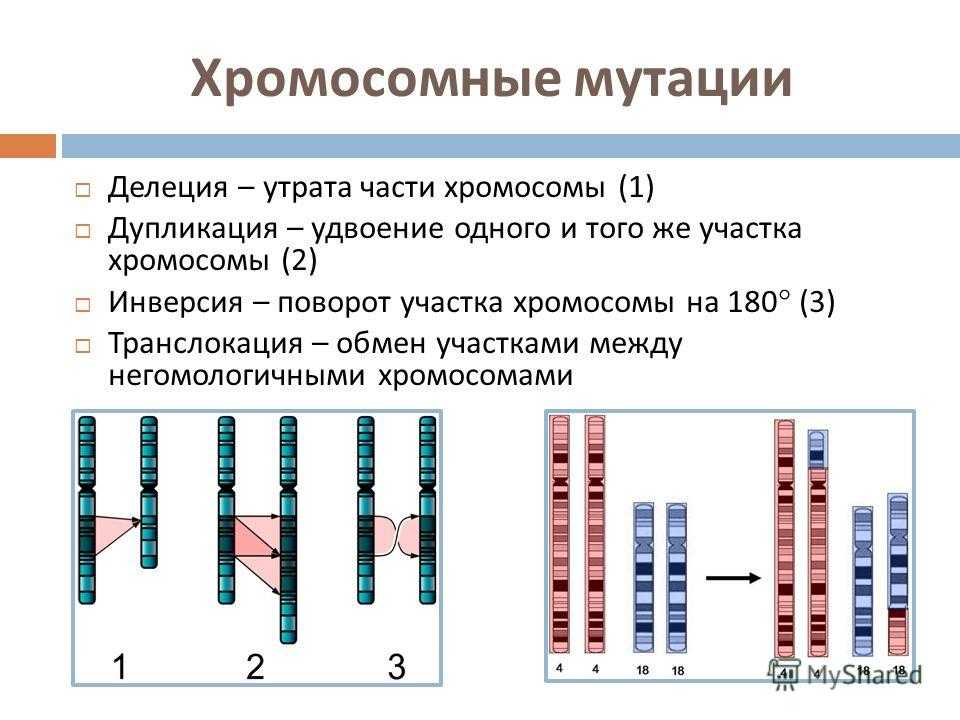
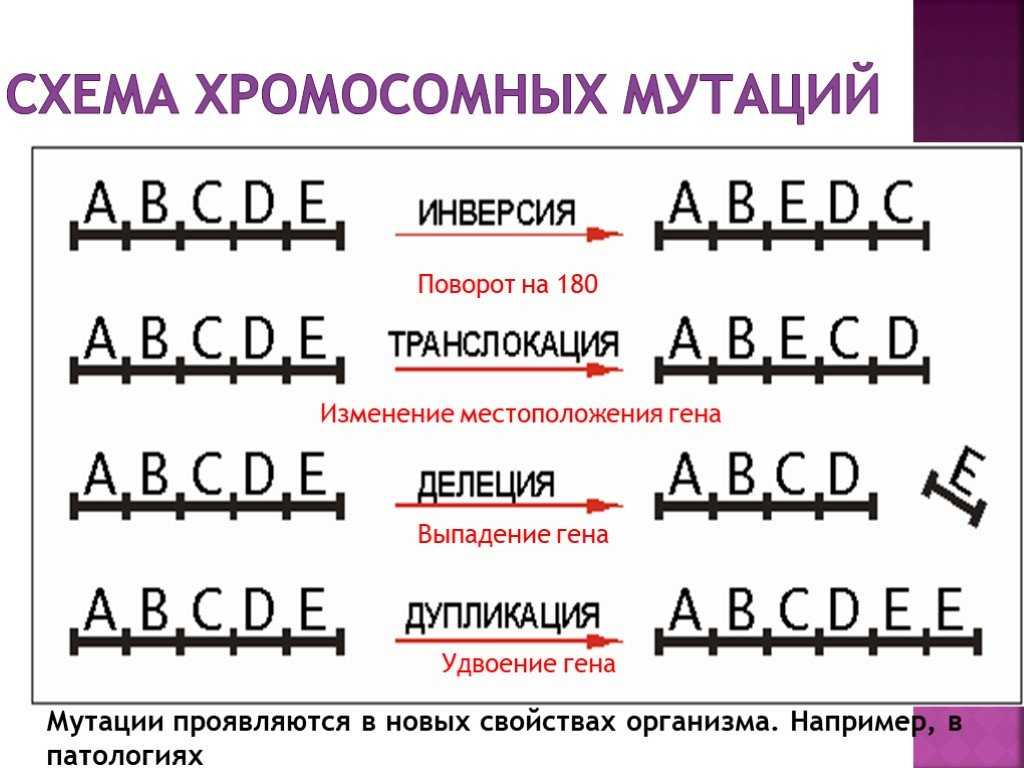
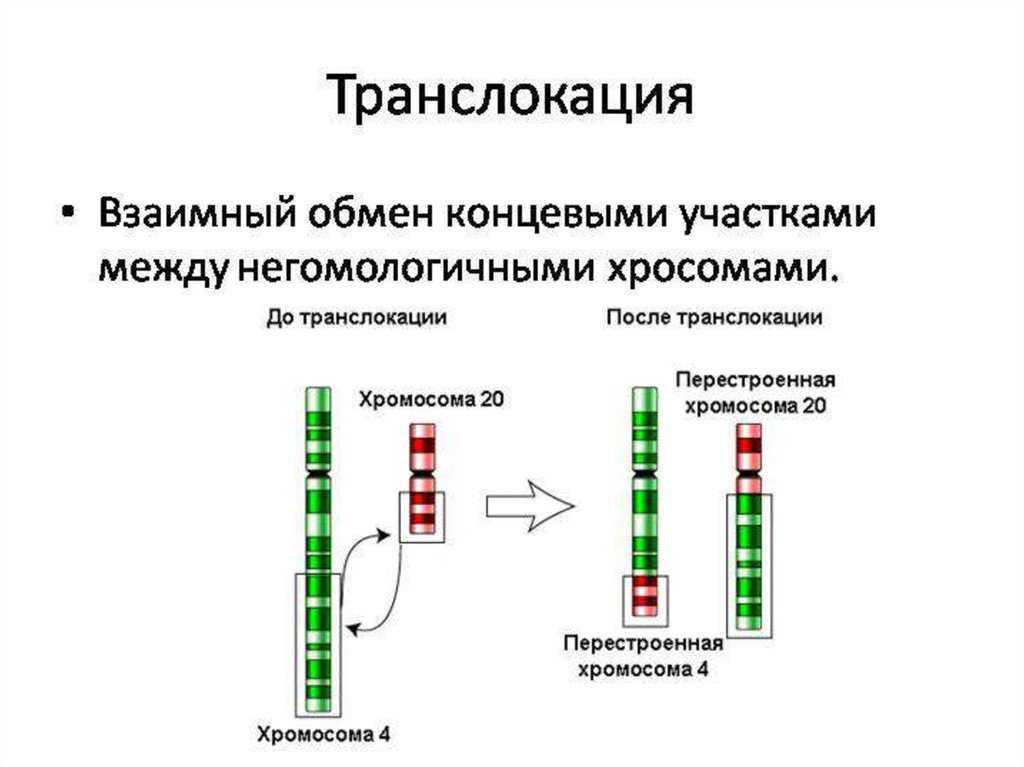
Робертсоновские транслокации
Акроцентрические хромосомы 13, 14, 15, 21 и 22 человека имеют очень короткие p-плечи, содержащие только хромосомные спутники и генетический код рибосомных РНК. Следовательно, их центромеры расположены очень близко к одному концу хромосомы. Когда длинные плечи двух акроцентрических хромосом сливаются в результате транслокации, их короткие плечи теряются и, в зависимости от расположения точек разрыва, создается дицентрическая или моноцентрическая гибридная хромосома.
Потеря коротких плеч акроцентрических хромосом не имеет фенотипических последствий, поскольку утраченные участки не содержат уникальных генетических последовательностей. Единственным следствием может быть сбалансированный кариотип с 45 хромосомами вместо 46. Это определяется примерно у 1 из 1000 человек.
Однако несбалансированные гаметы гетерозиготных носителей распространены и дают начало моносомному или трисомному плоду. Большинство моносомий и трисомий будут летальными и спонтанно прерываются на ранних сроках беременности. Однако выжившие плоды с трисомией 21 могут быть жизнеспособными и иметь синдром Дауна, хотя около 85% зачатых с трисомией 21 не доживают до срока. Когда этот синдром вызван родительской робертсоновской транслокацией, родитель-носитель обычно имеет слияние длинных плеч хромосом 14 и 21. Одна из трех жизнеспособных гамет будет сбалансирована, одна будет нормальной, а третья будет содержать слитую хромосому der(14;21), а также непораженную хромосому 21. Нормальное оплодотворение этой гаметы приводит к плоду с трисомией 21. Эмпирический риск для ребенка с синдромом Дауна составляет от 10 до 15 процентов для матери-носителя и только 2 процента, если отец — носитель робертсоновской транслокации.
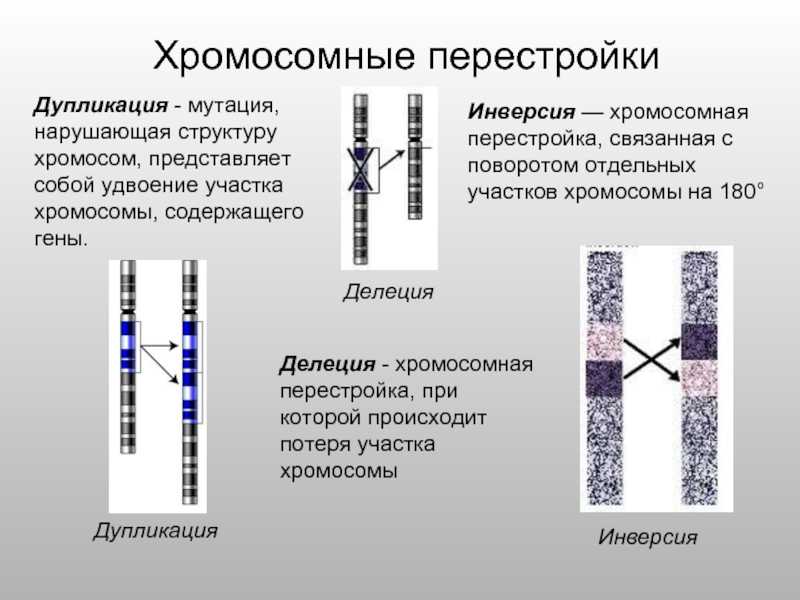
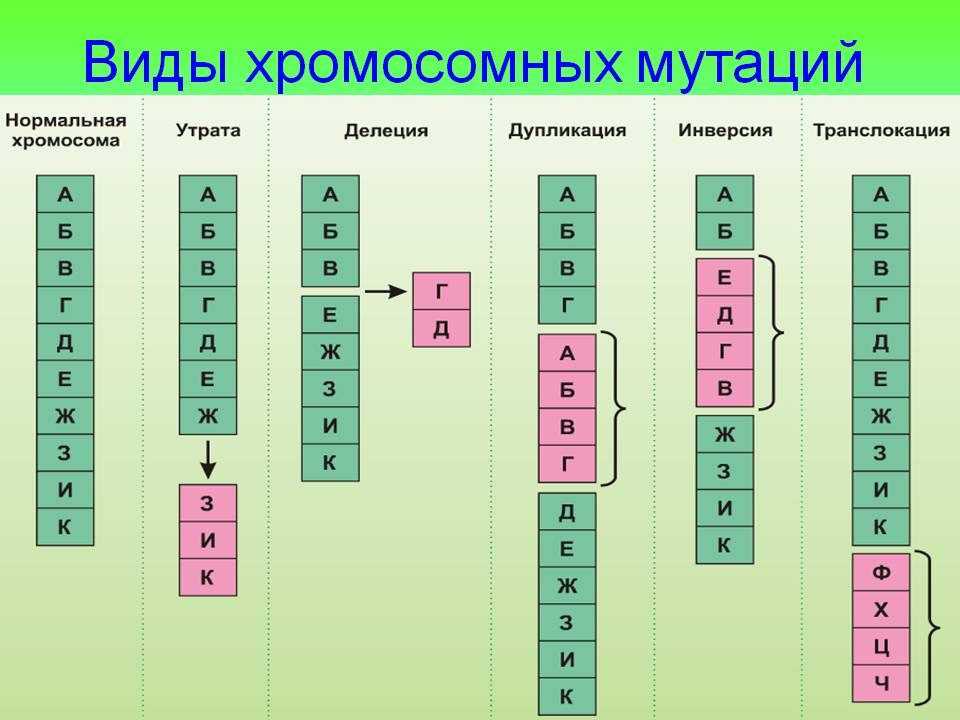
Дупликации[]
Файл:Gene-duplication-notext.png
Дупликация.
Дупликации появляются в результате неравного кроссинговера (в этом случае второй гомолог несет делецию) или в результате ошибки в ходе репликации. При конъюгации хромосомы с дупликацией и нормальной хромосомы как и при делеции формируется компенсационная петля.
Практически у всех организмов в норме наблюдается множественность генов, кодирующих рРНК (рибосомальную РНК). Это явление назвали избыточностью генов. Так у E. coli на рДНК (ДНК, кодирующее рРНК) приходится 0,4 % всего генома, что соответствует 5-10 копиям рибасомальных генов.
Другой пример дупликации — мутация Bar у Drosophila, обнаруженная в 20-х годах XX века Т. Морганом и А. Стертевантом. Мутация обусловлена дупликацией локуса 57.0 X-хромосомы. У нормальных самок (B+/B+) глаз имеет 800 фасеток, у гетерозиготных самок (B+/B) глаз имеет 350 фасеток, у гомозигот по мутации (B/B) — всего 70 фасеток. Обнаружены также самки с трижды повторенным геном — double Bar (BD/B+).
В 1970 году Сусумо Оно в монографии «Эволюция путем дупликации генов» разработал гипотезу об эволюционной роли дупликаций, поставляющих новые гены, не затрагивая при этом функций исходных генов. В пользу этой идеи говорит близость ряда генов по нуклеотидному составу, кодирующих разные продукты. Это трипсин и хемотрипсин, гемоглобин и миоглобин и ряд других белков.
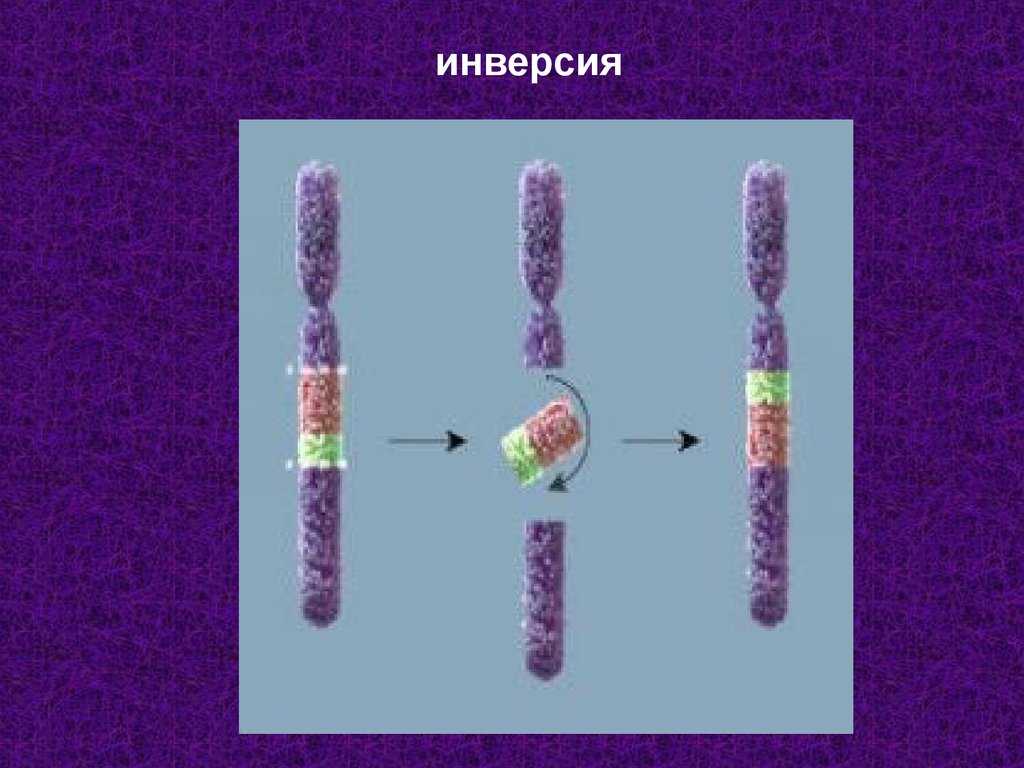
Что такое инверсия в биологии
Инверсия в биологии – это генетическое явление, характеризующееся перестановкой частей генетического материала внутри хромосомы. При инверсии происходит обратное расположение генетической последовательности. Такое изменение может возникнуть в результате ошибки в процессе мейоза или извне, под воздействием мутагенов или радиации.
Инверсия может быть перевернутой (парная инверсия) или неперевернутой (беспарная инверсия). Парная инверсия – это обратное расположение двух фрагментов одной и той же хромосомы. Беспарная инверсия – это переворот генетической последовательности, но без образования пары хромосом.
Во время инверсии могут возникать проблемы во время паринга хромосом в процессе мейоза. Это может привести к генетическим изменениям, таким как нарушение процесса рекомбинации или мутации.
Инверсии являются одной из форм структурных изменений хромосом и могут оказывать влияние на эволюцию и адаптацию организмов. Они могут приводить к изменению выражения генов, работе ферментов или протеканию процессов развития.
Примером инверсии является инверсия ног у насекомых. Обычно насекомые имеют прямоходящую ноги, но иногда наблюдаются популяции, у которых ноги перевернуты в обратную сторону. Это явление является примером парной инверсии и может быть связано с генетическими мутациями, которые произошли в процессе эволюции.
Что такое инверсия в генетике?
Инверсия в генетике — это тип структурной перестройки хромосом, при котором порядок генов внутри хромосомы меняется. В отличие от делеции (удаления генов) или дупликации (удвоения генов), при инверсии гены не теряются и не удваиваются, они просто меняют свою ориентацию на хромосоме.
Процесс инверсии происходит, когда две точки хромосомы обрываются и перекрепляются, меняя порядок генов между ними. Инверсии могут быть парными (одновременно происходят на обеих однонитях хромосом одной пары) или непарными (происходят только на одной из однонитей пары хромосом).
Инверсии в генетике могут быть видимыми или невидимыми, в зависимости от того, как они влияют на фенотип организма. Если инверсия не влияет на функционирование генов или не вызывает изменений в фенотипе, она считается невидимой. Видимые инверсии могут быть ассоциированы с различными генетическими нарушениями или генетическими заболеваниями.
Инверсии могут возникать как случайные мутации в процессе эволюции или быть унаследованными от одного из родителей. Они могут быть обнаружены с помощью методов молекулярной генетики, таких как цитогенетический анализ или методы секвенирования ДНК.
Инверсии в генетике могут иметь различные последствия, включая возможность нарушения процессов мейоза (деления половых клеток) и формирования анеуплоидных (неправильного числа хромосом) гамет. Они также могут приводить к изменениям в генной экспрессии (активности генов) или провоцировать перестройки генома.

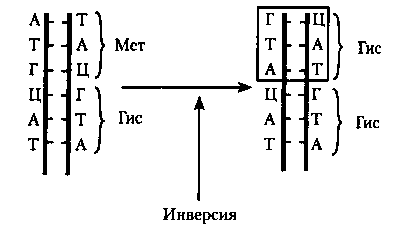
![5.1. мутации. генетика человека с основами общей генетики [учебное пособие]](https://lesniepolyani.ru/wp-content/uploads/a/e/d/aede9be6c6a2e399c13ad24d40c79369.jpeg)
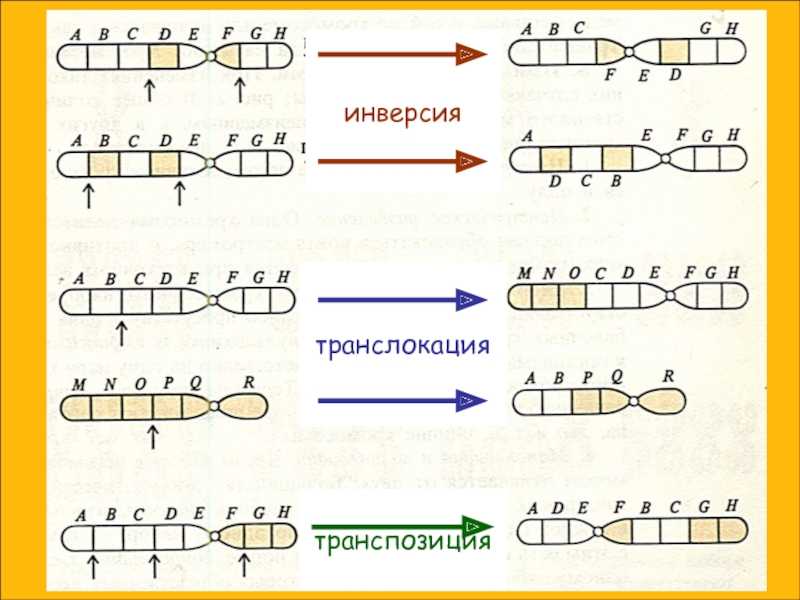
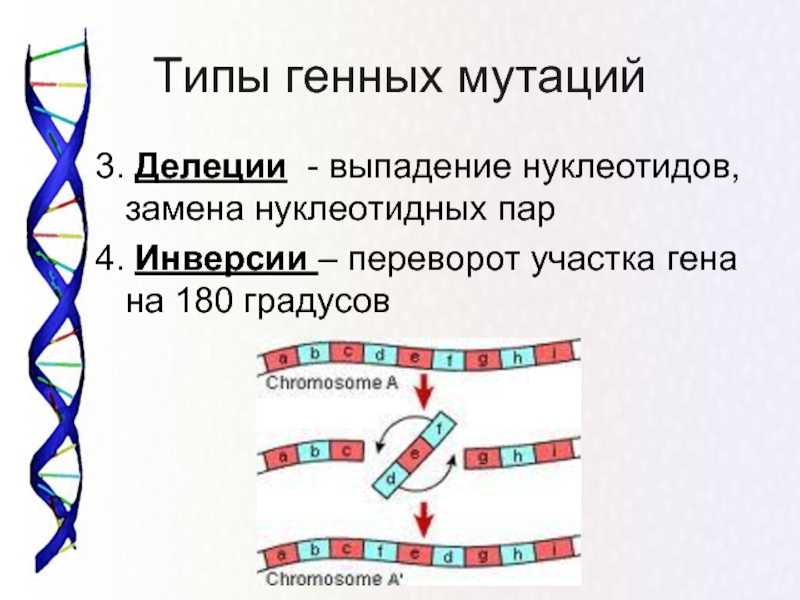
Выводы: инверсия в генетике — это структурная перестройка хромосом, при которой меняется порядок генов. Они могут быть видимыми или невидимыми и влиять на фенотип организма. Инверсии могут быть унаследованы или возникать как случайные мутации. Они могут иметь различные последствия, связанные с нарушением мейоза или изменением генной экспрессии.
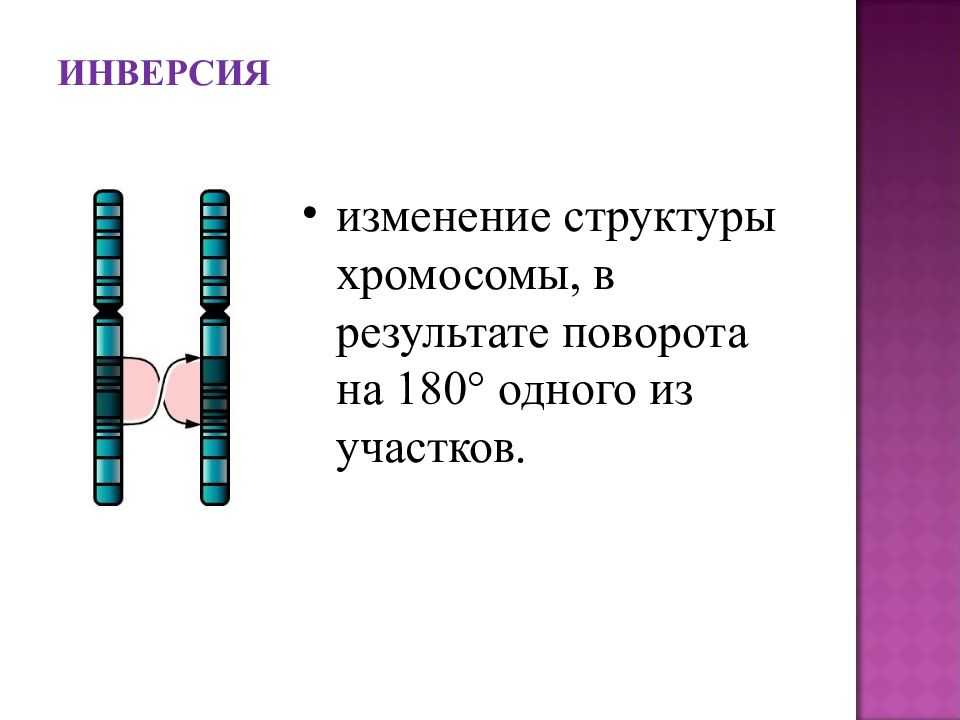
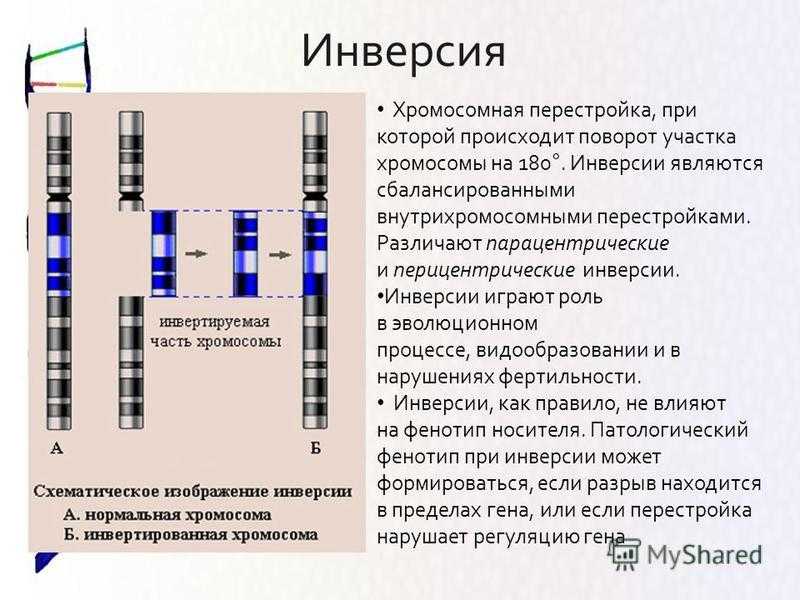
Инверсии[]
Файл:Chromosome inversion.png
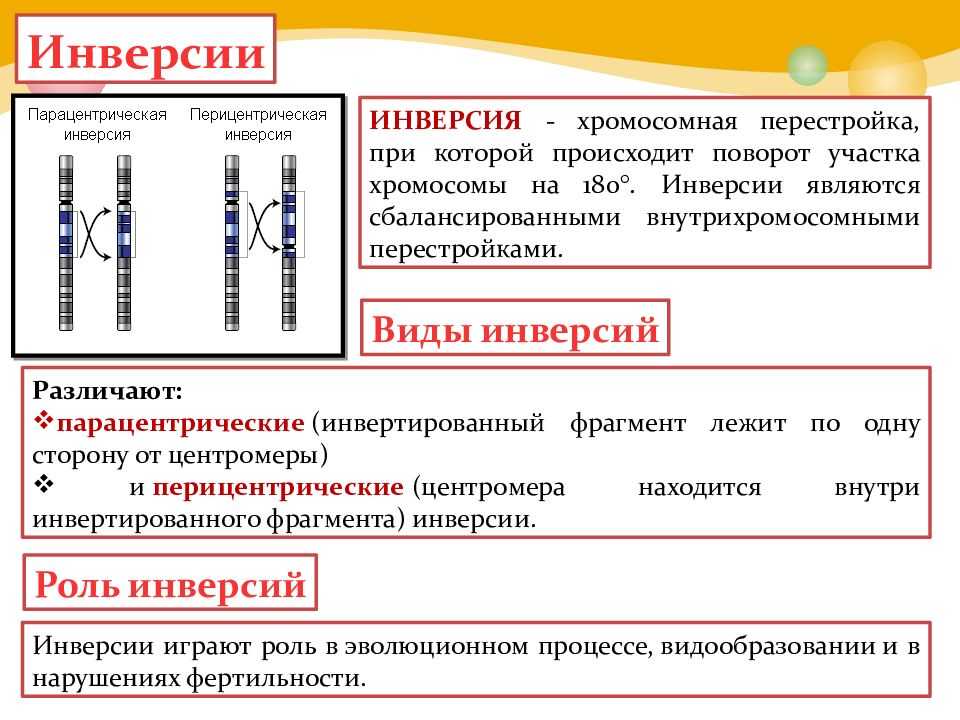
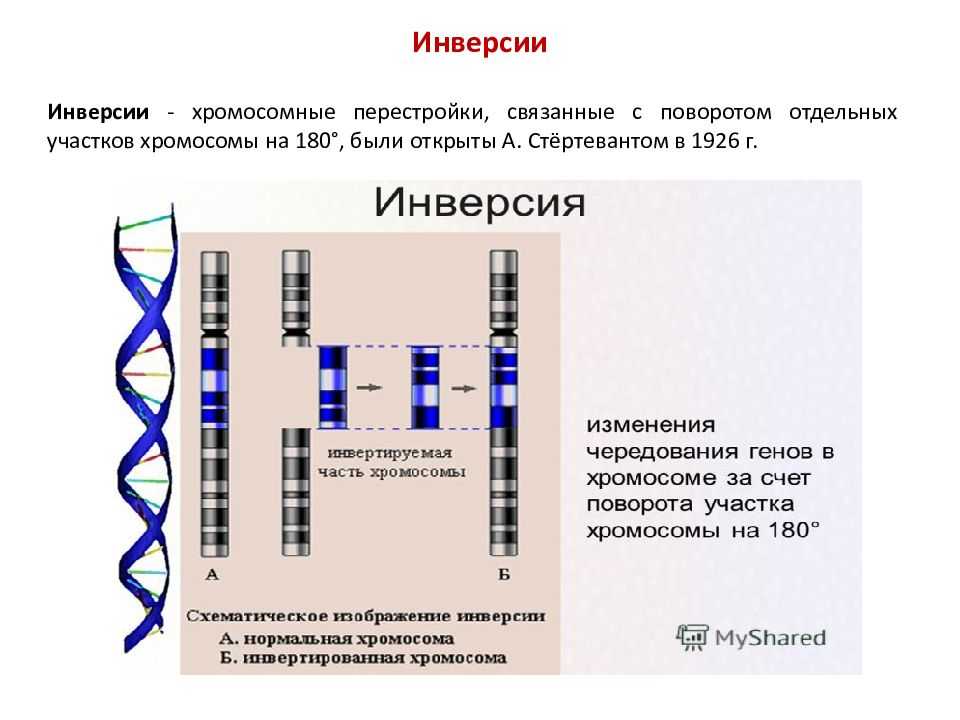
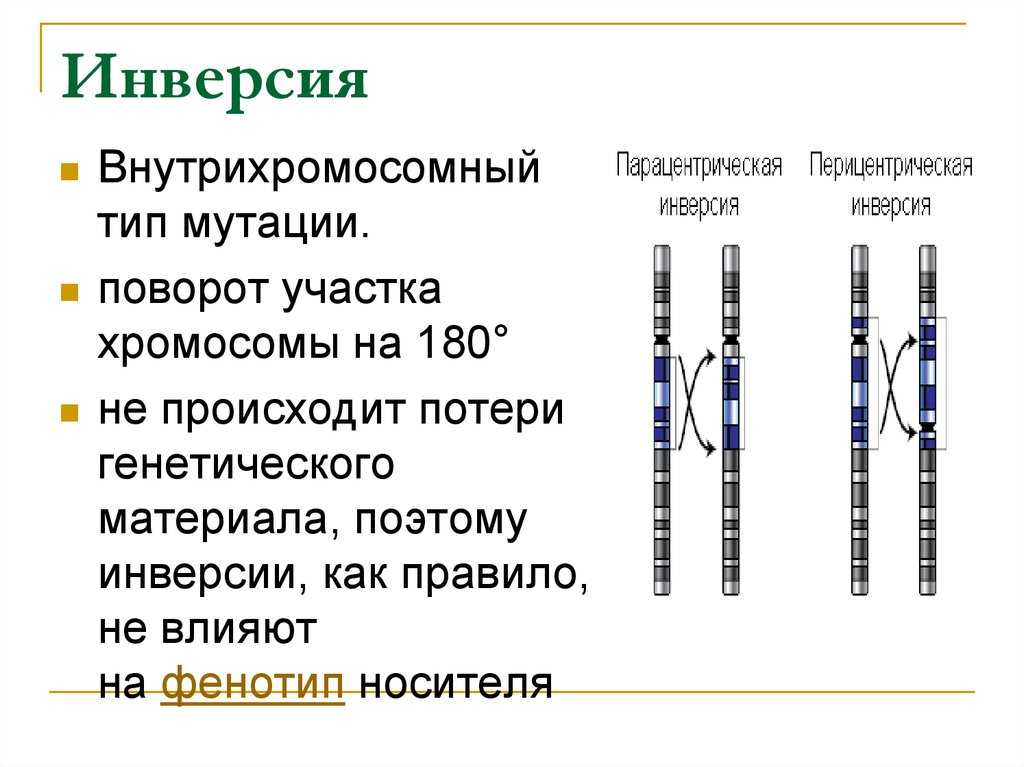
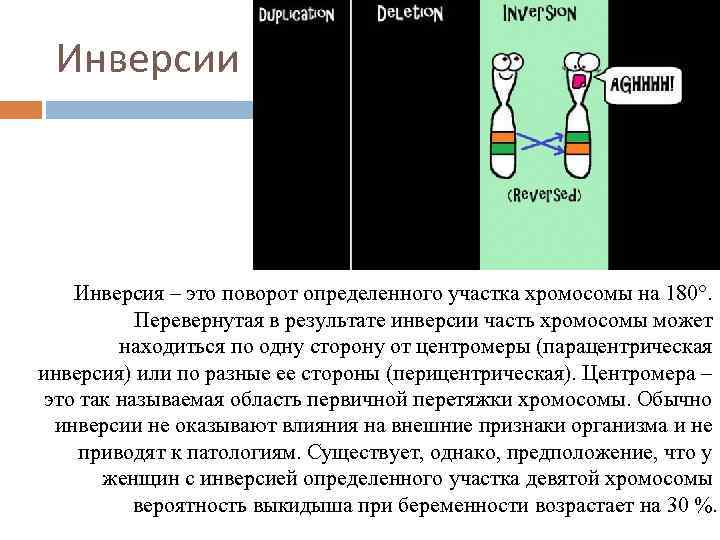
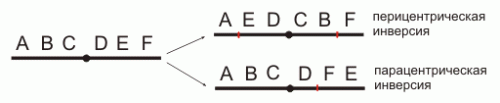
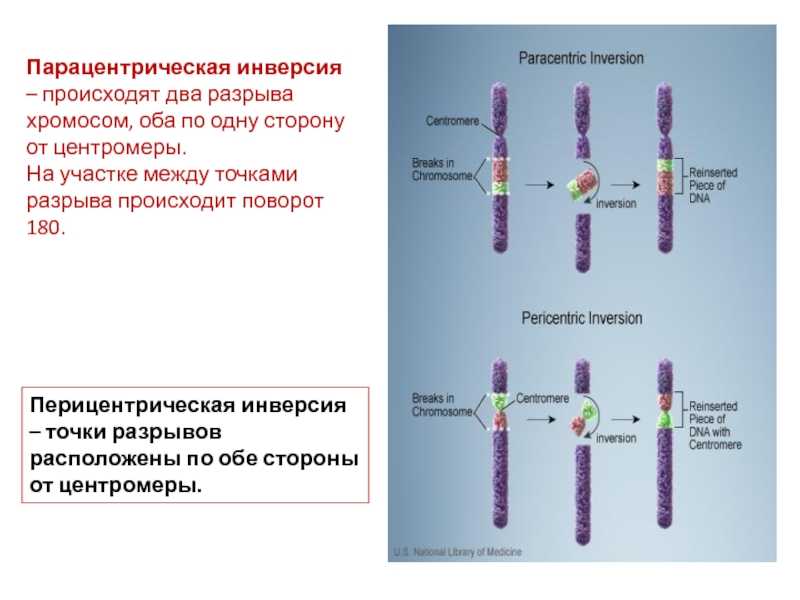
Инверсия. a — нормальная хромосома, b — парацентрическая инверсия, c — перицентрическая инверсия.
Различают парацентрические (инвертированный фрагмент лежит по одну сторону от центромеры) и перицентрические (инвертированный фрагмент лежит по разные стороны от центромеры) инверсии. При инверсиях не происходит потери генетического материала, потому как таковые инверсии как правило не влияют на фенотип, но если в инверсионной гетерозиготе (то есть организме, несущем как нормальную хромосому, так и хромосому с инверсией) происходит кроссинговер, то существует вероятность формирования аномальных хроматид. В случае парацентрической инверсии образуется одна нормальная и одна инвертированная (фенотипически нормальная) хроматиды, дицентрическая хроматида с дупликацией и делецией (при расхождении хроматид она обычно разрывается на две) и ацентрическая хроматида с дупликацией и делецией (обычно утрачивается). В случае перицентрической инверсии образуется одна нормальная и одна инвертированная хроматиды, а также две хроматиды с дупликацией и делецией. Гаметы, несущие дефектные хромосомы, обычно не развиваются или погибают на ранних этапах эмбриогенеза. Но гаметы с инвертированной хромосомой развиваются в организмы, 50 % гамет которых нежизнеспособны. Т.о. мутация сохраняется в популяции.
У человека наиболее распространенной является инверсия в 9 хромосоме, не вредящая носителю, хотя существуют данные, что у женщин с этой мутацией существует 30 % вероятность выкидыша.
Выявление инверсий
В настоящее время существует три основных подхода для выявления инверсий: с помощью классического генетического анализа, цитологически и на основе данных секвенирования полного генома . Наиболее распространённым является цитологический подход.
Именно при помощи генетического анализа инверсии были впервые обнаружены: в 1921 году Альфред Стёртевант показал инвертированный порядок идентичных генов у Drosophila simulans
по сравнению с Drosophila melanogaster
. Наличие инверсии можно предположить, если в скрещиваниях обнаруживается нерекомбинирующая часть генома, для этого метода необходимо предварительное генетическое картирование признаков.
Цитологически инверсии впервые наблюдали на политенных хромосомах слюнных желез у дрозофил, и двукрылые по-прежнему являются наиболее удобным объектом для наблюдения инверсий. В других таксономических группах крупные инверсии можно выявить при помощи дифференциальной окраски метафазных хромосом. Известные полиморфные варианты инверсий можно анализировать при помощи флуоресцентной гибридизации in situ
с использованием локус-специфических ДНК-проб.
У людей и у других видов с секвенированным геномом субмикроскопические инверсии можно обнаружить при помощи парноконцевого секвенирования . Межвидовые различия по инверсиям можно выявлять при помощи прямого сравнения гомологичных последовательностей .
Примечания
-
Бородин П. М., Торгашева А. А.
Хромосомные инверсии в клетке и эволюции // Природа. — 2011. — № 1
. — С. 19-26
. -
Генетика человека по Фогелю и Мотулски / М. Р. Спейчер, С. Е. Антонаракис, А. Г. Мотулски. — 4-е издание. — СПб: Н-Л. — С. 165-168. — 1056 с. — ISBN 978-5-94869-167-1 .
-
Kirkpatrick M.
How and why chromosome inversions evolve (англ.)
// PLoS biologу. — 2010. — Vol. 8, no. 9
. — P. e1000501. — PMID 20927412 . -
Sturtevant A.H.
A case of rearrangement of genes in Drosophila (англ.)
// Proc Natl Acad Sci USA. — Vol. 7, no. 8
. — P. 235-237. — PMID 16576597 . -
Korbel J. O. et al.
Paired-end mapping reveals extensive structural variation in the human genome (англ.)
// Science. — Vol. 318, no. 5849
. — P. 420-426. — PMID 17901297 . -
Feuk L. et al.
Discovery of human inversion polymorphisms by comparative analysis of human and chimpanzee DNA sequence assemblies (англ.)
// PLoS genetics. — 2005. — Vol. 1, no. 4
. — P. e56. — PMID 16254605 . -
Pfeiffer P., Goedecke W., Obe G.
Mechanisms of DNA double-strand break repair and their potential to induce chromosomal aberrations (англ.)
// Mutagenesis. — 2000. — Vol. 15, no. 4
. — P. 289-302. — PMID 10887207 . - Dittwald P. et al.
Inverted Low‐Copy Repeats and Genome Instability-A Genome‐Wide Analysis (англ.)
// Human mutation. — Vol. 34, no. 1
. — P. 210-220. —
Причины возникновения инверсии в геноме
1. Гомологичная рекомбинация
Одной из основных причин возникновения инверсии в геноме является гомологичная рекомбинация. В процессе этого биологического механизма две хромосомы с одинаковой последовательностью ДНК (гомологичные хромосомы) обмениваются участками ДНК. При неправильной ориентации и связывании этих участков может произойти инверсия, то есть изменение порядка генов.
2. Мобильные генетические элементы
Мобильные генетические элементы, такие как транспозоны и ретротранспозоны, могут также способствовать возникновению инверсии в геноме. Эти элементы способны перемещаться внутри генома и вставляться в различные места. При этом они могут вызывать инверсии, переставляя и меняя порядок генов на хромосоме.
3. Воздействие физических факторов
Некоторые физические факторы, такие как ионизирующее излучение и химические мутагены, могут повреждать ДНК и возбуждать рекомбинацию. В результате повреждения могут образовываться аберрации, включая инверсии в геноме.
4. Ошибки при репликации ДНК
При репликации ДНК могут возникать ошибки, что может приводить к образованию инверсий. Такие ошибки могут происходить в результате несовместимой связи между нуклеотидами, неправильной ориентации генетического материала или воздействия мутагенов. Результатом таких ошибок может быть образование инверсий в геноме.
5. Генетические перестройки
Генетические перестройки, такие как транслокации и делеции, могут также вызывать инверсии в геноме. При транслокации часть гена может переместиться на другую хромосому, а при делеции может пропасть часть гена. В результате таких перестроек могут возникать инверсии в геноме.



Некоторые причины возникновения инверсии в геноме
Причина
Описание
Гомологичная рекомбинация
Обмен участками ДНК между гомологичными хромосомами
Мобильные генетические элементы
Перемещение и вставка генетических элементов
Воздействие физических факторов
Ионизирующее излучение и химические мутагены
Ошибки при репликации ДНК
Неправильные связи и ориентация нуклеотидов
Генетические перестройки
Транслокации и делеции генов
Связь генных, геномных и хромосомных мутаций
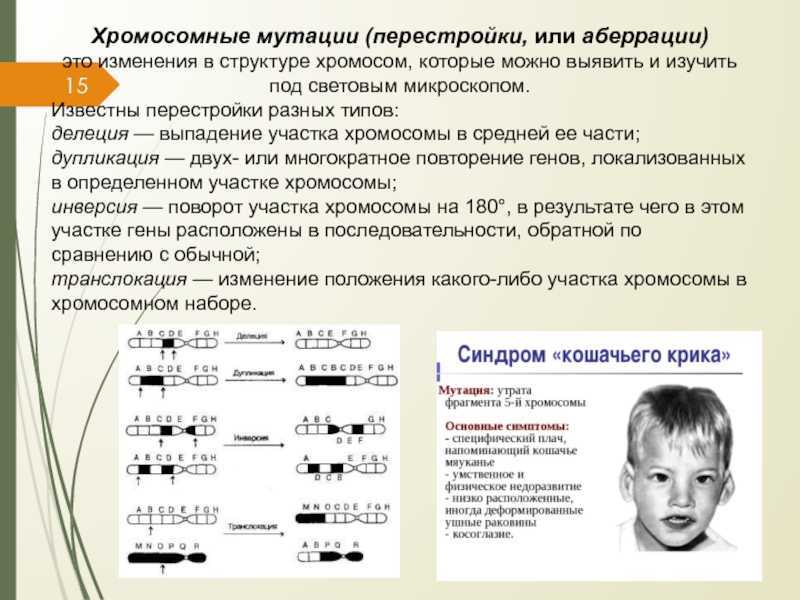
Являясь одним из видов наследственной изменчивости, мутационные изменения подразделяются на три взаимосвязанные разновидности: генные, геномные и хромосомные. Генные мутации происходят вследствие изменения строения генов, которые являются составной единицей хромосом. Геномные аберрации связаны с изменением числа хромосом, а хромосомные – с изменением их строения.
Геномные и хромосомные перестройки объединены общим медицинским понятием «хромосомные аномалии» (которые включают хромосомные болезни, внутриутбробные и соматические патологии) и общей классификацией в делении на анеуплодии и полиплодии. Центрические слияния (или робертсоновские транслокации) являются переходной формой от хромосомной перестройки к геномной. Треть из всех известных хромосомных аномалий описана в виде синдромов (групп патологий со схожими симптоматическими признаками).
Как инверсия влияет на организм
Инверсия — это генетическое явление, которое может повлиять на организм различными способами. Вот несколько основных способов, которыми инверсия может влиять на организм:
- Изменение структуры хромосом. Инверсия может привести к изменению порядка генов на хромосоме. Это может привести к возникновению новых характеристик или изменению уже существующих в организме.
- Нарушение процесса рекомбинации. Рекомбинация — это процесс обмена генетической информацией между хромосомами при производстве половых клеток. Инверсия может нарушить этот процесс, что может привести к ошибкам в распределении генов между потомками.
- Повышение риска возникновения генетических заболеваний. Инверсия может увеличить риск возникновения генетических заболеваний, особенно если соответствующий ген находится в области инверсии. Это связано с возможным нарушением экспрессии гена или изменением его функций.
- Обмен генетическим материалом с другими организмами. В некоторых случаях, инверсия может способствовать обмену генетическим материалом между организмами. Это может способствовать разнообразию и эволюции.
В целом, инверсия может иметь разное влияние на организм в зависимости от места и масштаба инверсии, а также от контекста, в котором она происходит. Более детальные исследования позволят лучше понять механизмы влияния инверсии на организмы и их эволюцию.
Делеции
Делеция хромосомного материала может быть обнаружена или не обнаружена стандартными методами хромосомного бэндинга (т. е. макроделеции и микроделеции соответственно).
Макроделяции — чтобы под микроскопом увидеть хромосомную делецию с помощью группировки хромосом, делеция должна охватывать не менее трех-пяти мегабаз (Mb). Учитывая, что средняя плотность генов составляет примерно один ген на 50 килобаз, неудивительно, что потеря такой большой области может привести к фенотипическим проявлениям. Возможно, наиболее известный пример — синдром кошачьего крика, возникающий из-за терминальной делеции короткого плеча хромосомы 5. Врожденные аномалии при этом расстройстве включают кошачий неонатальный плач, микроцефалию с дисморфическими чертами лица, пороки сердца, гипотонию и тяжелую умственную отсталость. Несмотря на то, что размер удаленного фрагмента различается от пациента к пациенту, всегда включаются хромосомные полосы 5p15.2 и 5p15.3. Таким образом, эти полосы охватывают критическую область для этого фенотипа.
Микроделеции — цитогенетические разработки, такие как методы выделения полос с высоким разрешением и флуоресцентная гибридизация in situ (FISH), позволили обнаружить относительно небольшие («микро») делеции. Если зонд FISH предназначен для локализации области, делетированной на одном аллеле, при оценке (про)метафазной хромосомы будет присутствовать только один двойной сигнал.
Один из примеров микроделеции — синдром Смита-Магениса, встречающийся у 1 к 25 000 и характеризующийся микроделецией участка хромосомы 17p11.2. Фенотипические особенности включают умственную отсталость, низкий и хриплый голос, агрессивное поведение, бессонницу, низкий рост и брахидактилию. В то время как большинство делеций охватывают около 5 Мб и цитогенетически различимы вблизи центромеры, остальные диагностируются с помощью более точных диагностических методов, таких как FISH. Интерстициальная делеция при этом синдроме облегчается наличием областей малокопийных повторов, фланкирующих общие области точек разрыва. Эти сегменты в высокой степени гомологичны, что делает их склонными к неравному кроссинговеру в мейозе I. Во время неравного кроссинговера две несестринские хроматиды рекомбинируют в неправильном месте из-за неправильного выравнивания высоко гомологичных последовательностей ДНК. Таким образом, одна из хроматид приобретает дупликацию промежуточной последовательности, в то время как соответствующий сегмент удаляется на другой хроматиде. Это называется неаллельной гомологичной рекомбинацией (NAHR) и лежит в основе большинства синдромов микроделеции/микродупликации.
Гомологическая рекомбинация во время митоза бывает редкой и аномальной. Тем не менее, неравный кроссинговер также может происходить при митотическом делении и приводит к неравному обмену сестринскими хроматидами. Основной механизм смещения между повторяющимися последовательностями такой же, как и в мейозе. Однако это аномальное событие в митозе приводит к соматической мутации и может предрасполагать к раку, если удаленная последовательность содержит (или будет частью) гена-супрессора опухоли.
Подгруппа пациентов с гиперэозинофильным синдромом имеет новое слияние FIP1L1-PDGFRA из-за большой интерстициальной делеции на хромосоме 4q12. Это приводит к слитому белку с конститутивной тирозинкиназной активностью, эозинофильной пролиферацией и повреждением органов-мишеней. Пациенты с этим слиянием часто реагируют на ингибитор тирозинкиназы, иматиниба мезилат. Диагноз этого слитого белка можно поставить, продемонстрировав потерю гена CHIC2 в удаленной области с помощью FISH или с помощью ПЦР, демонстрирующей наличие слитого транскрипта.
Полиморфизм по инверсиям у человека
Долгое время инверсии можно было выявлять только при помощи анализа G-бэндированных метафазных хромосом. Этот метод позволяет обнаруживать только крупные инверсии, при этом даже крупные инверсии при G-бэндинге могут остаться незамеченными из-за локального сходства рисунка G-полос. Классический цитогенетический анализ на основе дифференциальной окраски хромосом позволил обнаружить несколько полиморфных инверсий, распространённых в человеческой популяции и не имеющих клинического значения. Инверсии являются наиболее частым хромосомным полиморфизмом, выявляемыми в цитогенетических лабораториях, и самыми частыми из них являются перицентрические инверсии, детектируемые в гетерохроматиновых районах хромосом 1, 2, 3, 5, 9, 10 и 16. Например, известно, что более 1 % человеческой популяции являются носителями перицентрической инверсии в 9 хромосоме inv(9)(p12;q13), которую считают вариантом нормы . Наиболее частой инверсией, который включает эухроматин , является инверсия inv(2)(p11;q23), которая также считается нейтральной . Есть и другие более редкие варианты полиморфных инверсий, которые детектируются в отдельных группах и которые ведут свою историю от одного предкового мутационного события. Например, такая инверсия inv(10)(q11.22;q21.1) обнаруживается с частотой 0,11 % в Швеции .
Современные методы анализа генома, включающие парноконцевое секвенирование , сравнительный анализ геномов близкородственных видов, анализ неравновесного сцепления однонуклеотидных полиморфизмов (SNP), позволили выявить около 500 субмикроскопических полиморфных инверсий. Среди них, например, инверсия на хромосоме 8 (8p23.1) размером около 4.5 млн п.о, которая обнаруживается у 25 % здоровых людей .
Сдвиг рамки считывания
К генным относятся мутации обусловленные сдвигом рамки считывания, когда происходит изменение количества нуклеотидных пар в составе гена. Это может быть как выпадение, так и вставка одной или нескольких нуклеотидных пар в ДНК. Генных мутаций по типу сдвига рамки считывания больше всего. Наиболее часто они возникают в повторяющихся нуклеотидных последовательностях.
Вставка или выпадение нуклеотидных пар может произойти в следствие воздействия определенных химических веществ, которые деформируют двойную спираль ДНК.
Рентгеновское облучение может приводить к выпадению, т. е. делеции, участка с большим количеством пар нуклеотидов.
Вставки нередки при включении в нуклеотидную последовательность так называемых подвижных генетических элементов, которые могут менять свое положение.
К генным мутациям приводит неравный кроссинговер. Чаще всего он происходит в тех участках хромосом, где локализуются несколько копий одного и того же гена. При этом кроссинговер происходит так, что в одной хромосоме возникает делеция участка. Этот участок переносится на гомологичную хромосому, в которой возникает дупликация участка гена.
Если происходит делеция или вставка числа нуклеотидов не кратного трем, то рамка считывания сдвигается, и трансляция генетического кода зачастую обессмысливается. Кроме того, может возникнуть нонсенс-триплет.
![5.1. мутации. генетика человека с основами общей генетики [учебное пособие]](https://lesniepolyani.ru/wp-content/uploads/7/d/4/7d45bf8fb040711ef0af5df94d69d942.jpeg)

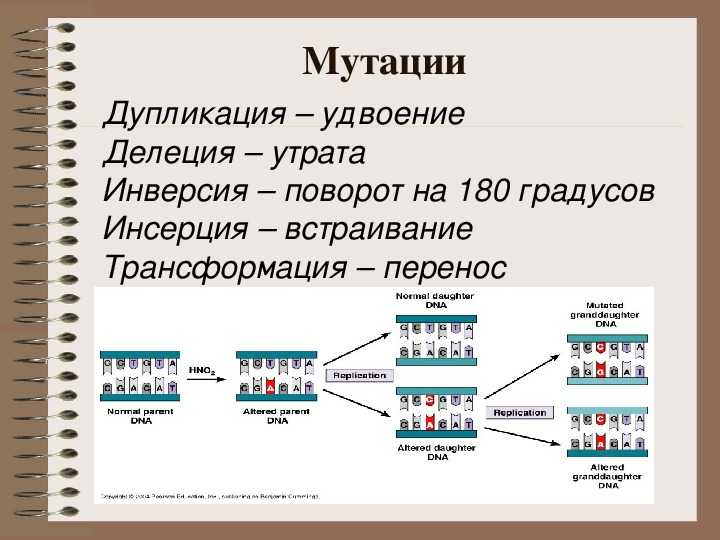

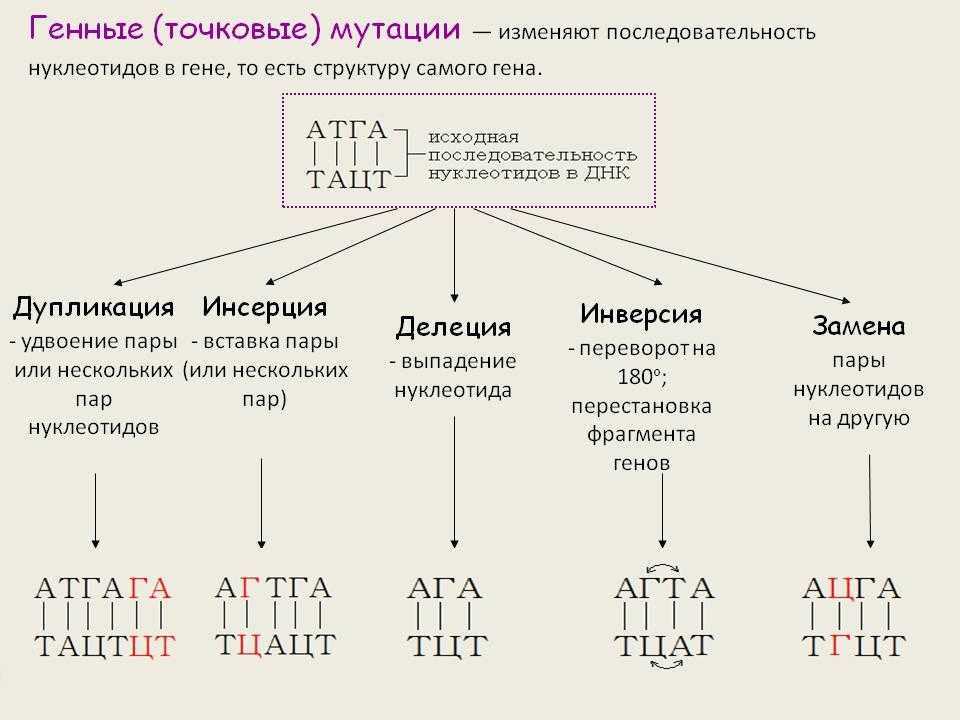

Если количество вставленных или выпавших нуклеотидов кратно трем, то, можно сказать, сдвиг рамки считывания не происходит. Однако при трансляции таких генов в пептидную цепь будут включены лишние или утрачены значащие аминокислоты.
Последствия инверсии в генетике
Инверсия является одной из самых распространенных структурных вариаций в геноме организмов. Она может иметь различные последствия для организма, в зависимости от расположения инверсии и ее размера.
В некоторых случаях инверсия не оказывает видимых последствий и организм сохраняет прежние фенотипические характеристики. Однако в других случаях инверсия может иметь значительные эффекты на организм.
Одно из последствий инверсии – возникновение генетических нарушений. В некоторых случаях инверсия может привести к изменению экспрессии генов или вызвать нарушение их функции. Это может привести к различным заболеваниям или нарушениям развития организма.
Инверсия также может оказывать влияние на процессы мейоза, в результате чего происходит неправильное распределение генетического материала между гаметами. Это может привести к формированию гамет с недостаточным или избыточным количеством генов, что может вызывать генетические нарушения у потомства.
Кроме того, инверсия может оказывать влияние на процесс рекомбинации. Рекомбинация – это процесс, в результате которого обменяющиеся генетическим материалом хромосомы образуют новые комбинации аллелей. Инверсия может изменить расположение генов на хромосомах и, следовательно, влиять на возможность образования новых комбинаций аллелей во время рекомбинации.
Инверсии также могут влиять на взаимодействие генов и эволюционные процессы. Изменение расположения генов на хромосомах может привести к изменению их связи и взаимодействия. Такие изменения могут оказать значительное влияние на эволюцию организма и способствовать возникновению новых фенотипических особенностей.
В целом, инверсия – это сложный процесс, который может иметь разнообразные последствия для организма. Точные эффекты зависят от конкретной инверсии и ее влияния на генетический материал и процессы в организме. Изучение этих последствий позволяет нам лучше понять генетические основы различных заболеваний и эволюции организмов.