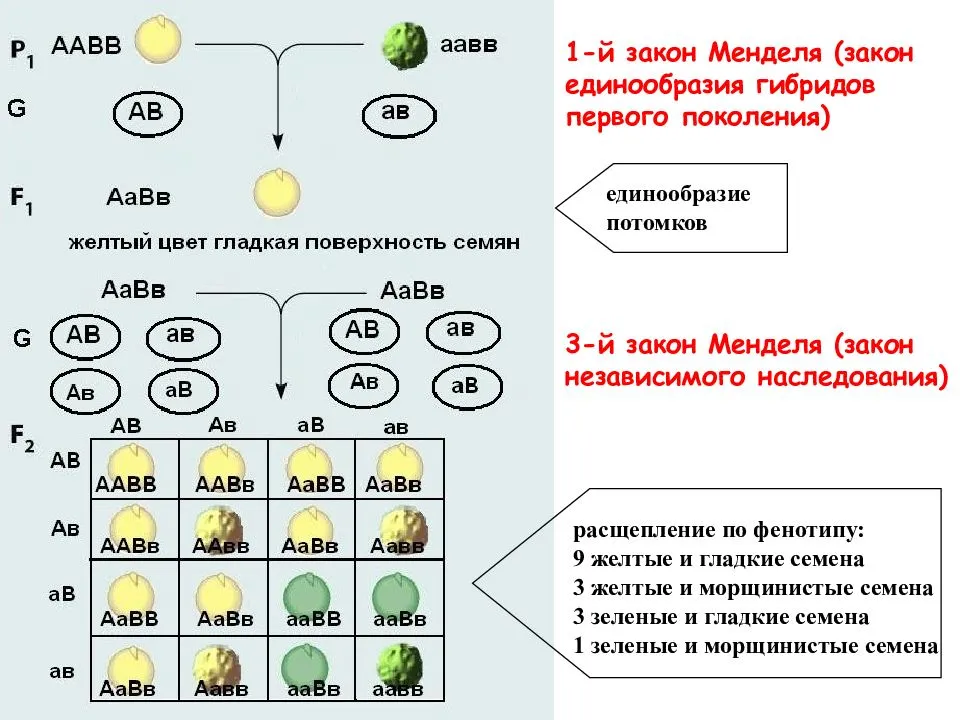
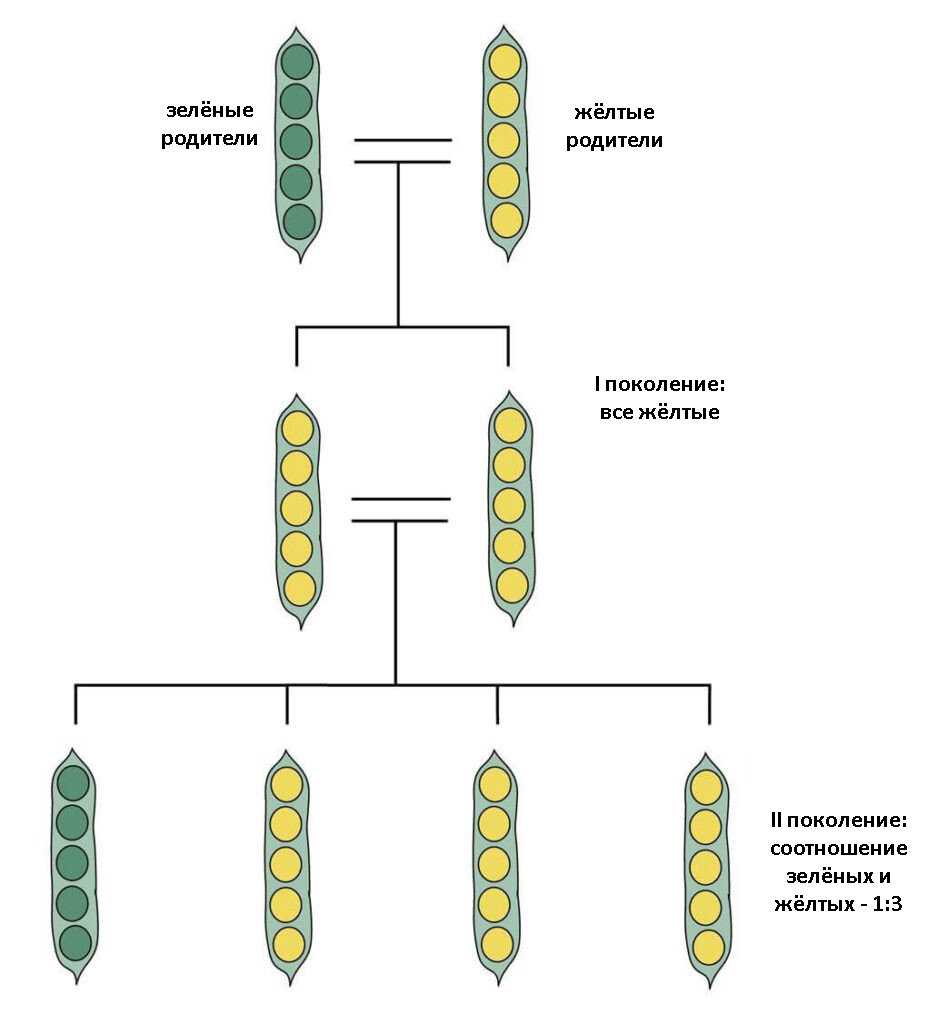
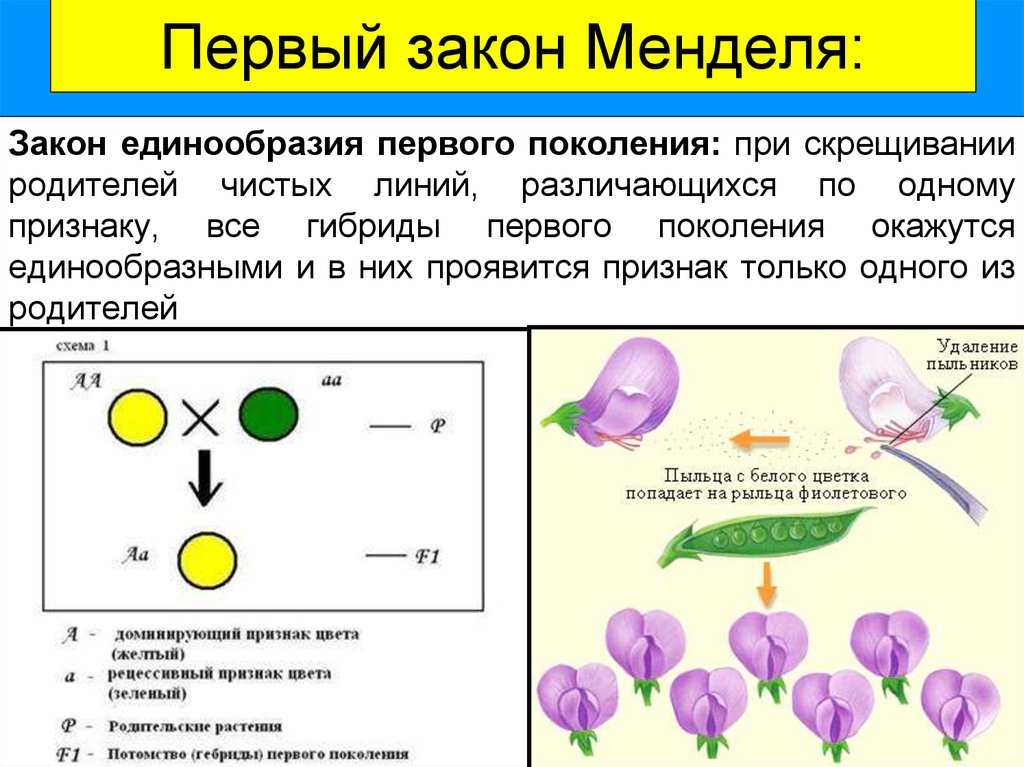
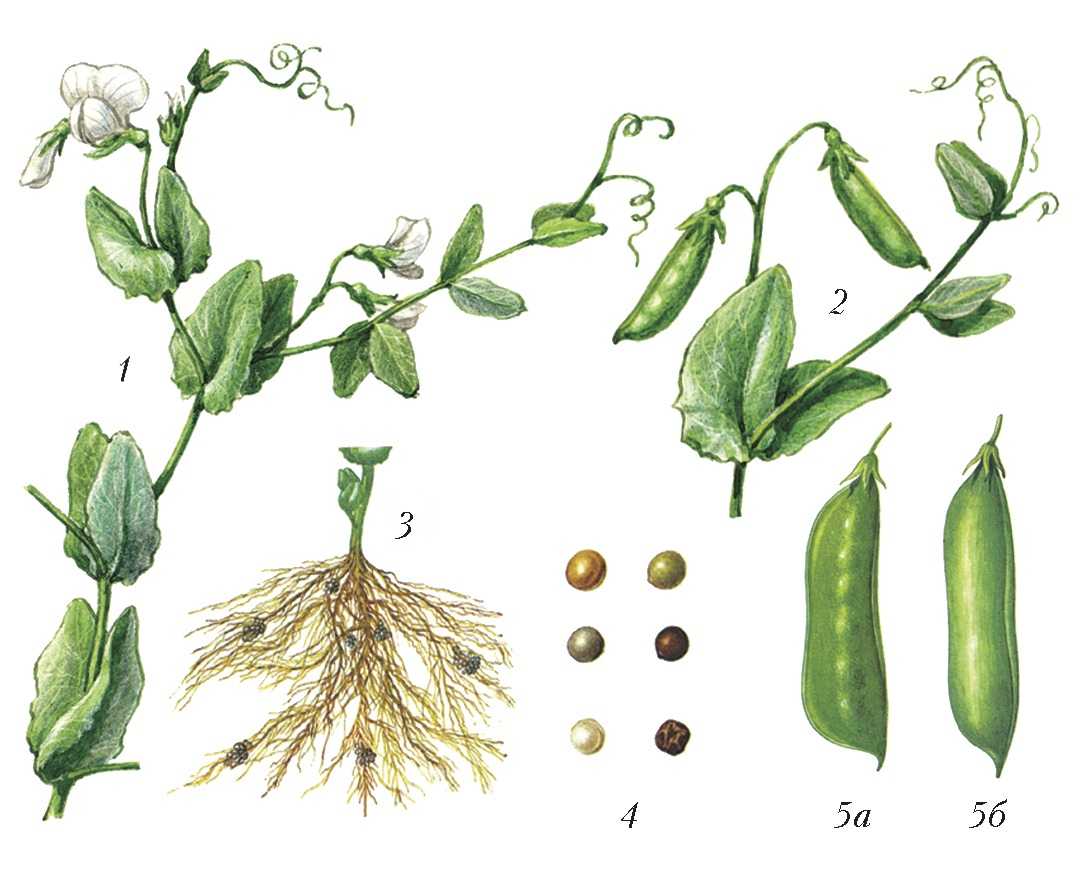
Генотипи зі зміненим морфотипом стебла
Стебло гороху округле, нечітко чотирьохгранне, всередині порожнисте, легко вилягає. Довжина стебла в залежності від сорту та умов вирощування вар’ює від 25 до 250—300 см. За формою стебло може бути просте, коли в межах його продуктивної частини вузли розташовані більш менш рівномірно, та фасційоване, коли стебло у верхній продуктивній частині плоско розширене, вузли зближені, квітки та боби розташовані зкучено, що нагадує суцвіття зонтик. Завдяки компактному розташуванню квіток і бобів, подальше цвітіння та дозрівання фасційованих рослин гороху проходить майже одночасно, але дещо пізніше, ніж у звичайних сортів.
Генотипи із детермінантним типом росту (ДТР)
ДТР сприяє формуванню генеративних органів тільки у верхній частині стебла та характеризується тим, що точка росту закінчується генеративною брунькою, на відміну від звичайних сортів, у яких ріст стебла продовжується навіть при формуванні на ньому бобів, що знижує можливість використання фотоасимилятів на утворення бобів, тим самим знижуючи насіннєву продуктивність рослини. В умовах достатнього, а особливо надмірного зволоження, необмежений ріст призводить до стовбуріння стебла. Надмірне нарощування вегетативної маси викликає вилягання рослин у порівняно ранніх фазах розвитку (до завершення наливу бобів). На нижніх плодоносах боби виявляються недостатньо виповненими, з сильною череззерницею, що різко знижує насіннєву продуктивність рослин, а в підсумку — врожай насіння. Ці недоліки в основному відсутні у рослин, які мають детермінантний (обмежений) ріст стебла.
Вперше форма гороху з обмеженим типом росту та термінальним розташуванням бобів (детер) була отримана Поповою І.А. в 1972 році в результаті обробки насіння сорту Свобода 10 нитрозоетилсечовиною (НЕС) в концентрації 0,012%, на основі якої був виведений сорт Атол з обмеженим типом росту. W.K. Swiecicki в 1987 році описав одержану ним лінію гороху з обмеженим ростом, WT 16100, констатував ідентичність її з лінією JI 1238 з інституту Джона Інеса, ввів термін det (determinate) для позначення гена, який контролює ознаку „обмежений ріст“ та локалізував його в VII хромосомі: tl-det = 18.68; det-r = 18.7. Коротко цей тип можна описати наступним чином: на рослині утворюється один-два плодючі вузли, з останнього вузла виходять два плодоноси, точка росту закінчується генеративною брунькою.
Детери були залучені в селекційні програми в якості батьківських форм. А.М. Шевченко створів перший сорт Детермінантний ВСГІ, який поєднав гени def (неосипання насіння), af (безлисточковість) і det (детермінантний тип росту).
У Всеросійському науково-дослідному інституті зернобобових та круп’яних культур (Росія) в селекційний процес були залучені й інші нові морфотипи: „хамелеони”, які характеризуються ярусною гетерофілією та люпиноїди, які характеризуються детермінантним типом росту із формуванням апікального суцвіття. З селекційної точки зору „хамелеони” привертають увагу тим, що не зменшуючи стійкості до вилягання, вони мають більшу асиміляційну поверхню листків у порівнянні з безлисточковими формами. У 2006 р. У Державне сортовипробування в Україні були включени перші сорти типу ”хамелеон”. Люпиноїди, в свою чергу, привертають увагу обмеженим типом росту та багатоплідністю, в останньому вузлі немає ні вегетативної бруньки, ні листка, рослина повністю закінчує свій ріст після формування одного-двох плодючих вузлів.
Короткостеблові генотипи гороху
Короткостеблові сорти гороху, в порівнянні із звичайними сортами, характеризуються вищим рівнем стійкості до вилягання. Так, у низькорослих сортів із міцним стеблом рослини починають вилягати лише під час цвітіння, тоді як у високорослих сортів із довгим стеблом — ще до початку бутонізації (у фазу 8—10 листків). Короткостеблові сорти гороху характеризуються добре розвиненою механічною тканиною, мають розвинену транспортну систему стебла та краще забезпечення асимилятами генеративних органів. У короткостеблових сортів доля бобів в сухій масі рослин у фазі воскової стиглості насіння складає 40—50%, стебло — 30—40%, тоді як у довгостеблових сортів більша частина біомаси припадає на стебло (50—60%), а менша — на боби (12—20%).
Генеалогический анализ коллекционных сортообразцов гороха гетерофильной формы
Основная часть нашей работы посвящена проблеме, которая является центральной в селекции сельскохозяйственных культур, в том числе и гороха, — изучению исходного материала с целью подбора родительских пар для гибридизации.
Н.И. Вавилов (1935) для выявления наиболее результативных способов подбора родительских пар для скрещиваний применял генеалогический анализ. Это позволило сделать вывод о преимуществе гибридизации эколого и географически отдаленных форм.
Объектом наших исследований явилась коллекция, состоящая из 36 сор-тообразцов гороха морфотипа хамелеон. Ее изучение было начато с анализа родословной коллекционных сортообразцов, который позволил выявить основные способы ведения селекционной работы и особенности генетической детерминации признаков.
Как показал генеалогический анализ, всем сортообразцам в процессе селекции передавался ген 1т, контролирующий укороченность междоузлий. Растение гетерофильной формы с таким геном представлено на рисунке 4.1.1. Введение гена укороченности междоузлий в генотип гетерофильной формы проводилось с целью увеличения уборочного индекса, который является критерием использования биологического потенциала. Помимо увеличения уборочного индекса, уменьшение длины стебля в сочетании с усатыми листьями, каковые имеются в среднем ярусе гетерофильной формы, способствует также повышению устойчивости к полеганию.
С целью дальнейшего сокращения вегетативной массы в пользу семенной продуктивности предпринята попытка создания сортов с редуцированными прилистниками. В изучаемой коллекции имеется линия с редуцированными прилистниками Аз-123 (рис. 4.1.2).Признак редукции прилистников контролируется геном st. Линия Лз-123 была получена от скрещивания донора этого признака гетерофильной лини Б-104 с усатым сортом Ус-89-1798.
Традиционный морфотип гороха унаследовал от своих предков адаптированный к кустарниковому фитоценозу нелимитированный рост стебля и способность формировать почти неограниченное число продуктивных узлов (Макашева, 1973, 1979). В условиях достаточной влажности стебель израста-ет, образуя дополнительные продуктивные узлы, на которых генеративные органы могут гибнуть на стадии цветка, либо, сформировав бобы, не образовывать полноценных семян. На все это идут затраты невосполнимой энергии и элементов питания. При израстании растений их стеблестой становится более склонными к полеганию. Поэтому культурное растение должно обладать фиксированным ростом.
Последнее обстоятельство явилось причиной для работы над созданием гетерофильных сортов с детерминантным типом роста стебля. Коллекционным сортообразцам, передавался ген deh, контролирующий самарскую модель детерминантности стебля (рис. 4.1.3.)
В настоящее время коллекция насчитывает 9 гетерофильных сортообраз-цов с детерминантным типом роста стебля (самарская модель): Орел, Аз-26, Аз-376, Аз-378, Аз-1210, Аз-1211, Аз-1229, Аз-1233, Аз-1397.
В процессе селекции коллекционным сортообразцам передавался признак наличия прицвстничков. В результате большинство гетерофильных сортооб-разцов унаследовали небольшие одиночные прицветнички, контролируемые аллелями br, bra. Сортообразцам Орел, Аз-26, Аз-135, Аз-176 были переданы крупные парные прицветнички, обусловленные аллелем brae.
Неосыпаемость семян, контролируемая геном def, передана сортообразцам: Орел, Аз-122, Аз-130, Аз-176, Аз-325, Аз-365, Аз-367, Аз-376, Аз-378, Аз-468, Аз-1189, Аз-1397, Аз-1420, Аз-1433, Аз-1565.
Все перечисленные выше признаки свободно сочетаются с ярусной гете-рофиллией. При их передаче соответствующим сортообразцам использовались необходимые доноры. Отбор проводили из расщепляющихся гибридных комбинаций F2…F7. Доноры, несущие передаваемый признак, были в основном листочковых и усатых морфотипов, поэтому основной тип скрещиваний межморфный (так назван нами тип скрещиваний, когда хамелеон скрещивается с другими морфотипами) с использованием простой и сложи осту пенча-той гибридизации.
Скрещивание хамелеонов с донорами других морфотипов позволяет значительно увеличивать генетическое многообразие пребридинговой коллекции, что имеет большое значение в селекционной работе.
Селекція гороху на стійкість до хвороб і шкідників
У гороху є реальним використання наступних способів підвищення і збільшення тривалості стійкості до патогенів:
- разове використання генів стійкості;
- створення багатолінійних сортів;
- інтеграція різних олігогенів стійкості в одному генотипі;
- посилення горизонтальної стійкості при трансгресивній селекції;
- посилення толерантності до патогенів;
- створення сортів із груповою стійкістю до різних патогенів;
- використання генної інженерії для інтрогресії чужорідних генів стійкості.
Разове використання генів стійкості (РВГС)
Для реалізації селекційної програми РВГС потрібна наявність чітко позначених донорів моногенної стійкості до патогену, тобто потрібен сильний ген або блок генів, що стабільно виявляє свою дію в різних грунтово-кліматичних умовах і забезпечує стійкість до декількох найбільш вірулентних рас паразита.
Найбільш швидкий і простий шлях інтрогресії гена стійкості в сприйнятливий сорт — створення стійких ізоліній даного сорту шляхом схрещувань, що насичують, тобто беккроссів. Для здійснення схеми беккросової селекції гороху при домінантному контролі стійкості потрібно 7 поколінь, при рецесивному — 8 поколінь.

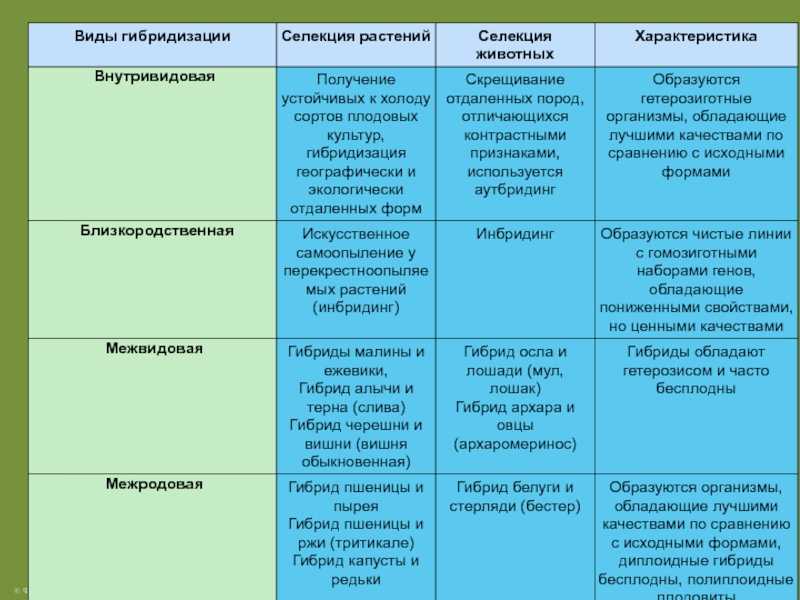
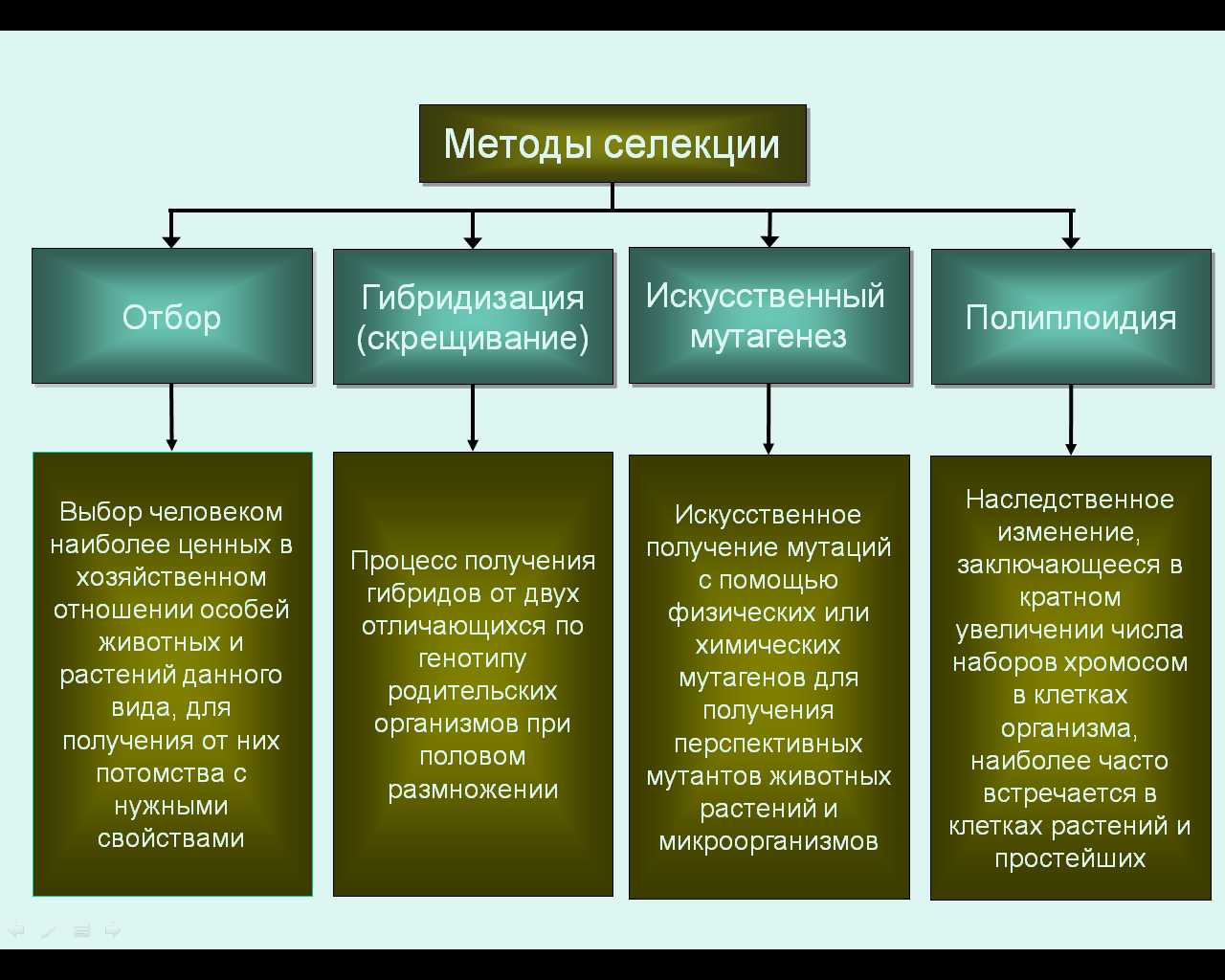



Схема схрещування виглядає в такий спосіб:
A(rr) — сприйнятливий сорт-реципієнт;
B(RR) — сорт-донор стійкого гена R;
1-e покоління (F1) — A(rr) x B(RR) = AB(Rr);
2-e покоління (F2B1)- AB(Rr) x A(rr) = 2AB(R-);
3-e покоління 2AB(R-) x A(rr) = 3АВ(R-);
4-e покоління 3AB(R-) x A(rr) = 4АВ(RR, Rr)
5—6-e покоління 4AB(RR, Rr) — самозапилення і добір стабільно стійкіх рослин із генотипом A(RR);
7-е покоління — розмноження і сортовипробування стійких ізоліній сорту А.
При використанні приміщень штучного клімату програма беккроссової селекції гороху по інтрогресії гена стійкості здійснюється за 2—3 роки при вирощуванні 3—4-х поколінь у рік. Метод РВГС у селекції гороху — найбільш ефективний по створенню сортів, стійких до факультативних сапрофітів, наприклад, до видів Fusarium.
Багатолінійна селекція
Для хвороб, які викликаються облігатними паразитами (пероноспороз, борошниста роса, іржа, вірусні хвороби), метод РВГС може бути недостатнім для підтримки тривалої специфічної стійкості — для них більше підходить варіант багатолінійної селекції. Для цього підбирають серію донорів стійкості до різних рас патогену і шляхом беккроссів створюють серію ізоліній високоврожайного сорту, що відрізняються друг за стійкістю до різних рас. Дослідження ряду авторів показують, що сорт, складений з декількох таких ліній, має досить високу і стабільну стійкість за рахунок стабілізації расового складу патогена шляхом зведення до мінімуму розвитку рас із множинним набором генів вірулентності. Слід зазначити, що стосовно облигатних паразитів неприпустимо тривале оброблення в одному регіоні одного або декількох сортів, специфічна стійкість яких контролюється тим самим олигогеном. Уповільнення розвитку нематоди і ґрунтові грибні паразити спостерігалися при використанні в посівах сумішей ліній.
Пірамідування різних генів стійкості
Вавилов особливу увагу звертав на створення сортів із груповим імунітетом, тобто стійкістю одночасно до комплексу фізіологічних рас або різних хвороб. Найкращим методом він вважав схрещування географічно віддалених форм культурних рослин і їхніх диких родичів, що відрізняються великою розмаїтістю генів стійкості.
- Може бути три різних випадки об’єднання генів в один генотип:
інтеграція (пірамідування) олигогенів, що контролюють специфічну стійкість до різних фізіологічних рас визначеного паразита;
інтеграція в одному сорті генів неспецифічної стійкості різного географічного походження до даної хвороби;
Учение Н. И. Вавилова о центрах происхождения и многообразия культурных растений
|
Н.И.Вавилов |
Селекция — наука о создании новых и улучшении существующих пород животных, сортов растений, штаммов микроорганизмов. В основе селекции лежат такие методы, как гибридизация и отбор. Теоретической основой селекции является генетика.
Для успешного решения задач, стоящих перед селекцией, академик Н.И.Вавилов особо выделял значение:
-
Изучения сортового, видового и родового разнообразия интересующей нас культуры;
-
Влияния среды на развитие интересующих селекционера признаков;
-
Изучения наследственной изменчивости;
-
Знаний закономерностей наследования признаков при гибридизации;
-
Особенностей селекционного процесса для само- или перекрестноопылителей;
-
Стратегии искусственного отбора.
Породы, сорта, штаммы — искусственно созданные человеком популяции организмов с наследственно закрепленными особенностями: продуктивностью, морфологическими, физиологическими признаками.
Каждая порода животных, сорт растений, штамм микроорганизмов приспособлены к определенным условиям, поэтому в каждой зоне нашей страны имеются специализированные сортоиспытательные станции и племенные хозяйства для сравнения и проверки новых сортов и пород.
Для успешной работы селекционеру необходимо сортовое разнообразие исходного материала, с этой целью Н.И.Вавиловым была собрана коллекция сортов культурных растений и их диких предков со всего земного шара. К 1940 году во Всесоюзном институте растениеводства насчитывалось 300 тыс. образцов. Но с позиций лысенковщины, занявшей в то время руководящие позиции в биологической науке России и считавшей, что определяющую роль в создании новых форм играет окружающая среда, эта коллекция была не нужна. Работы по пополнению коллекции были прекращены. В настоящее время коллекция пополняется и является основой для работ по селекции любой культуры.
Н.И.Вавилов установил центры происхождения культурных растений, где находится наибольшее видовое и сортовое многообразие культурных растений.
Центры происхождения культурных растений (по Н.И.Вавилову).
|
Центры происхождения |
Местоположение |
Культивируемые растения |
|
1. Южноазиатский тропический 2. Восточноазиатский 3. Юго-Западноазиат-ский 4. Средиземноморский 5. Абиссинский 6. Центральноамериканский 7. Южноамериканский |
Тропическая Индия, Индокитай, о-ва Юго-Восточной Азии Центральный и Восточный Китай, Япония, Корея, Тайвань Малая Азия, Средняя Азия, Иран, Афганистан, Юго-Западная Индия Страны по берегам Средиземного моря Абиссинское нагорье Африки Южная Мексика Западное побережье Южной Америки |
Рис, сахарный тростник, цитрусовые, баклажаны и др. (50% культурных растений) Соя, просо, гречиха, плодовые и овощные культуры — слива, вишня и др. (20% культурных растений) Пшеница, рожь, бобовые культуры, лен, конопля, репа, чеснок, виноград и др. (14% культурных растений) Капуста, сахарная свекла, маслины, клевер (11% культурных растений) Твердая пшеница, ячмень, кофейное дерево, бананы, сорго Кукуруза, какао, тыква, табак, хлопчатник Картофель, ананас, хинное дерево. |
Наиболее богатыми по количеству культур являются древние центры цивилизации, именно там наиболее ранняя культура земледелия, более длительное время проводится искусственный отбор и селекция растений.
Список литературы Применение математических методов анализа в селекции гороха овощного
- Хавалойес П. Зернобобовые. Питательные зерна устойчивого будущего. -ФАО, 2016. -196 с.
- Зотиков В.И., Наумкина Т.С., Грядунова Н.В. и др. Зернобобовые культуры -важный фактор устойчивого экологически ориентированного сельского хозяйства/Зернобобовые и крупяные культуры. -2016. -№ 1 (17). -С.6-13.
- Международный год зернобобовых 2016. . -Режим доступа: http//www.fao.org/pulses-2016/ru. -(Дата обращения: 02.02.2017).
- Казыдуб Н.Г., Кузьмина С.П. Селекционная работа с зернобобовыми культурами в Омском ГАУ/Зернобобовые культуры -развивающееся направление в России: материалы первого междунар. форума. -Омск: ОмГАУ, 2016. -С.5-9.
- Кузьмина С.П., Казыдуб Н.Г., Мерзлякова Е.В. и др. Хозяйственно-биологическая характеристика образцов коллекции гороха овощного в Омском ГАУ/Состояние и перспективы развития садоводства в Сибири: материалы II Нац. науч.-практ. конф., посвящ. 85-летию плодового сада Ом. ГАУ им. проф. А.Д. Кизюрина. -Омск, 2016. -С.69-72.
- Бондаренко Е.В., Кузьмина С.П. Морфобиологические особенности коллекционных образцов гороха овощного в условиях южной лесостепи Омской области/Роль аграрной науки в устойчивом развитии сельских территорий. -Новосибирск, 2017. -С.22-29.
- Вишнякова М.А., Булынцев С.В., Бурляева М.О. и др. Исходный материал для селекции овощных зернобобовых культур в коллекции ВИР/Овощи России. -2013. -№ 1 (18). -С.16-26.
- Самарин Н.А., Самарин С.Н. Возможные пути сохранения семеноводства отечественных сортов гороха овощного в современных условиях/Овощи России. -2013. -№ 1 (18). -С.76-78.
- Волощенко В.С. Пути интенсификации производства гороха овощного в России/Экономика с.-х. и перераб. предпр. -2011. -№1. -С.33-35.
- Государственный реестр селекционных достижений, допущенных к использованию. -М., 2016. -505 с.
- Корсаков Н.И., Адамова О.А., Будакова В.И. и др. Методические указания по изучению коллекции зерновых бобовых культур. -Л.: ВИР, 1975. -59 с.
- Бююль А. SPSS: искусство обработки информации. Анализ статистических данных и восстановление скрытых закономерностей. -СПб.: ДиаСофтЮП, 2002. -608 с.
- Кайгородова И.М., Пышная О.Н., Пронина Е.П. Изучение наследования скороспелости у гороха овощного/Овощи России. -2013. -№1 (18). -С.35-40.
- Зотиков В.И., Бударина Г.А. Болезни гороха и основные приемы защиты культуры в условиях средней полосы России/Защита и карантин растений. -2015. -№5. -С.11-15.
- Ушаков В.А., Пронина Е.П. Сортимент гороха овощного селекции ВНИИССОК/Овощи России. -2013. -№1. -С. 63-65.
- Зубов А.Е., Катюк А.И. Методы и результаты селекции гороха в Самарском НИИСХ/Изв. Самар. науч. центра РАН. -2014. -Т. 16, № 5-3. -С.1127-1130.
- Пивоваров Н.Ф., Цыганок Н.С. Восстановить производство сушеного зеленого горошка/Овощи России. -2011. -№1 (12). -С.43-48.
- Корнева С.П. Использование кластерного анализа для повышения эффективности отборов в расщепляющихся гибридных популяциях/Молодые ученые Сибирского региона -аграрной науке. -Омск, 2004. -Вып.4. -С.127-131.
Основные методы селекции животных
Создание пород домашних животных началось вслед за их приручением и одомашниванием, которое началось 10-12 тыс. лет назад. Содержание в неволе снижает действие стабилизирующей формы естественного отбора. Различные формы искусственного отбора (сначала бессознательный, а затем методический) приводят к созданию всего многообразия пород домашних животных.









В селекции животных, по сравнению с селекцией растений, есть ряд особенностей. Во-первых, для животных характерно в основном половое размножение, поэтому любая порода является сложной гетерозиготной системой. Оценка качеств самцов, которые внешне у них не проявляются (яйценоскость, жирномолочность), оцениваются по потомству и родословной. Во-вторых, у них часто поздняя половозрелость, смена поколений происходит через несколько лет. В-третьих, потомство немногочисленное.
Основными методами селекции животных являются гибридизация и отбор. Различают те же методы скрещивания — близкородственное скрещивание, инбридинг, и неродственное — аутбридинг. Инбридинг, как и у растений, приводит к депрессии. Отбор у животных проводится по экстерьеру (определенным параметрам внешнего строения), т.к. именно он является критерием породы.
1. Внутрипородное разведение направлено на сохранение и улучшение породы. Практически выражается в отборе лучших производителей, выбраковке особей, не отвечающих требованиям породы. В племенных хозяйствах ведутся племенные книги, отражающие родословную, экстерьер и продуктивность животных за много поколений.
2. Межпородное скрещивание используют для создания новой породы. При этом часто проводят близкородственное скрещивание, родителей скрещивают с потомством, братьев с сестрами, это помогает получить большее число особей, обладающих нужными свойствами. Инбридинг сопровождается жестким постоянным отбором, обычно получают несколько линий, затем производят скрещивание разных линий.
Хорошим примером может служить выведенная академиком М.Ф.Ивановым порода свиней — украинская белая степная. При создании этой породы использовались свиноматки местных украинских свиней с небольшой массой и невысоким качеством мяса и сала, но хорошо приспособленных к местным условиям. Самцами-производителями были хряки белой английской породы. Гибридное потомство вновь было скрещено с английскими хряками, в нескольких поколениях применялся инбридинг, были получены чистые линии, при скрещивании которых получены родоначальники новой породы, которые по качеству мяса и массе не отличались от английской породы, по выносливости — от украинских свиней.
3. Использование эффекта гетерозиса. Часто при межпородном скрещивании в первом поколении проявляется эффект гетерозиса, гетерозисные животные отличаются скороспелостью и повышенной мясной продуктивностью. Например, при скрещивании двух мясных пород кур получают гетерозисных бройлерных кур, при скрещивании беркширской и дюрокджерсейской пород свиней получают скороспелых свиней с большой массой и хорошим качеством мяса и сала.
4. Испытание по потомству проводят для подбора самцов, у которых не проявляются некоторые качества (молочность и жирномолочность быков, яйценоскость петухов). Для этого производителей-самцов скрещивают с несколькими самками, оценивают продуктивность и другие качества дочерей, сравнивая их с материнскими и со среднепородными.
5. Искусственное осеменение используют для получения потомства от лучших самцов производителей, тем более что половые клетки можно хранить при температуре жидкого азота любое время.
6. С помощью гормональной суперовуляции и трансплантации у выдающихся коров можно забирать десятки эмбрионов в год, а затем имплантировать их в других коров, эмбрионы так же хранятся при температуре жидкого азота. Это дает возможность увеличить в несколько раз число потомков от выдающихся производителей.
7. Отдаленная гибридизация, межвидовое скрещивание, известно с древних времен. Чаще всего межвидовые гибриды стерильны, у них нарушается мейоз, что приводит к нарушению гаметогенеза. С глубокой древности человек использует гибрид кобылицы с ослом — мула, который отличается выносливостью и долгожительством. Но иногда гаметогенез у отдаленных гибридов протекает нормально, что позволило получить новые ценные породы животных. Примером являются архаромериносы, которые, как и архары, могут пастись высоко в горах, а, как мериносы, дают хорошую шерсть. Получены плодовитые гибриды от скрещивания местного крупного рогатого скота с яками и зебу. При скрещивании белуги и стерляди получен плодовитый гибрид — бестер, хорька и норки — хонорик, продуктивен гибрид между карпом и карасем.