

Введение
В селекционных программах интересующие сорта и образцы часто выращивают и тестируют в нескольких локациях в течение нескольких лет. Такая серия испытаний называется испытаниями в нескольких локациях (multi-environment trial, МЕТ), где сочетание «год — локация» называется средой (environment). Чтобы количественно оценить и в итоге сравнить точность MET, селекционеры часто рассчитывают показатель наследуемости в узком смысле (narrow-sense heritability, h2) или наследуемости в широком смысле (broad-sense heritability, H2) на основе значения генотипа (genotype-mean basis). H2 определяется как доля фенотипической изменчивости, относящаяся к общей изменчивости генотипа, включая аддитивную, доминантную и эпистатическую изменчивость . Наследуемость является ключевым фактором для достижения высоких показателей генетического прогресса (genetic gain), поэтому понимание того, как использовать этот показатель в селекционных программах, необходимо для того, чтобы разрабатывать и выпускать сорта и гибриды, отвечающие потребностям сельхозтоваропроизводителей и потребителей.
Цели данного руководства:
- внести ясность в значение показателя наследуемости;
- показать, как можно рассчитать наследуемость, используя подходящие методы;
- предоставить рекомендации по надежным методам количественной оценки и сравнения точности полевых испытаний в селекционных программах.
Взаимодействие генотип-среда в процессе селекции
Выше была рассмотрена роль компонентов генетической изменчивости и соотношения генетической и экологической изменчивости в выраженности одного единственного признака. Однако как между отдельными признаками и факторами окружающей среды, так и между генотипом как единым целым (особенно в отношении урожая) и факторами окружающей среды может произойти взаимодействие (VGE), что следует учитывать в ходе процесса селекции.
Создание новых сортов растений — обычно процесс длительный, и селекционный материал подвергается действию факторов окружающей среды на протяжении большого числа лет. На создание и передачу в производство нового сорта однолетних растений в среднем уходит около 10 лет, для многолетних растений — значительно больше.
Начиная с F2, проводят отбор фенотипов, у которых, как ожидается, произойдет рекомбинация генов с проявлением положительных агрономических признаков. Вследствие сильного ежегодного варьирования условий среды один год может быть благоприятным для испытаний на засухоустойчивость, второй — для оценки на устойчивость к низким температурам, третий — для испытаний на устойчивость к болезням и т.д. После 5-6 лет отбора можно ожидать, что материал, выдержавший все эти испытания, обнаружит широкую приспособляемость и она предохранит его от возникновения отрицательного взаимодействия генотип — сезон года. У такого материала трудно, но можно ожидать положительного использования им большинства благоприятных факторов среды, причем совсем не обязательно его урожайность должна быть на самом высоком уровне. Кроме того, вполне возможно, что повторные испытания будут проходить в поколениях расщепления, когда значительная часть материала еще гетерозиготна. Позже, в процессе формирования линий, их отбор проводят даже при отсутствии низких температур, засухи или болезней; лишь при широком использовании этих линий в производстве они обнаруживают ранее неустановленные недостатки.
Поэтому, чтобы не зависеть от нерегулярности лимитирующих факторов окружающей среды, в процессе отбора принято создавать искусственные условия (используя теплицы, фитотроны, лаборатории) и материал в поколениях расщепления, а также первоначально отобранные растения и линии испытывать на устойчивость к низким температурам (в условиях Югославии до -15°С), на засухоустойчивость, устойчивость к болезням и т.д. Для более полной проверки влияния климата и патогенных организмов значительное число селекционных учреждений выращивает материал в расщепляющихся поколениях и проводит отбор по крайней мере в двух различных географических районах, что в немалой степени может заменить сезоны года. Все эти испытания снижают возможность возникновения риска от неблагоприятного взаимодействия генотип — окружающая среда.
О взаимодействии генотип — среда будет более подробно рассказано в главе о приспособляемости и стабильности сортов.
Влияние наследственности на интеллект и характер в разных возрастах
Исследования
показывают, что гены отвечают за 50-70%
разнообразия людей по уровню интеллекта
и за 28-49% различий по выраженности
пяти «универсальных», наиболее важных,
свойств личности:
- уверенности в себе,
- тревожности,
- дружелюбию,
- сознательности,
- интеллектуальной гибкости.
Это данные — для
взрослых. Однако степень влияния
наследственности зависит от возраста. Результаты психогенетических
исследований не подтверждают распространенное
мнение, что с возрастом гены все меньше
влияют на поведение человека. Генетические
различия, как правило, сильнее проявляются
в зрелом возрасте, когда характер уже
сформировался. Значения коэффициента
наследуемости большинства изученных
психологических свойств для взрослых
выше, чем для детей. Наиболее точные данные
получены по наследственной обусловленности
интеллекта. В младенчестве внутрипарное
сходство многояйцевых близнецов такое
же высокое, как для однояйцевых, но после
трех лет оно начинает снижаться, что можно
объяснить большим влиянием генетических
различий. При этом нарастание различий
происходит не линейно. В ходе развития
ребенка есть этапы, в которые различия
между детьми вызываются преимущественно
влиянием среды. Для интеллекта это возраст
3-4 года, а для формирования личности —
предподростковый возраст 8-11 лет.
Кроме того, в
разных возрастах действуют разные
генетические факторы. Так среди
наследственных факторов, обуславливающих различия
по интеллекту, есть как стабильные, т.е.
действующие во всех возрастах (это, возможно,
гены, связанные с так называемым «общим
интеллектом»), так и специфические
для каждого периода развития (вероятно,
гены, определяющие развитие частных способностей).
История исследования
От чего зависит
поведение человека? Что влияет на
характер и темперамент, на интеллект
и творчество – наследственность или воспитание?
Спор об этом ведется уже сотни лет. Одно
из первых научных исследований этого
вопроса принадлежит английскому биологу
Фрэнсису Гальтону, двоюродному брату
Чарльза Дарвина. В 1865 г. он опубликовал
статью “Наследственный талант и характер”,
а позже еще одну работу -“История близнецов
как критерий относительной силы природы
и воспитания”.
Гальтон сравнил
близнецов, которые были “очень похожи
при рождении” с теми, которые были похожи
как обычные братья и сестры. Теперь таких
близнецов называют монозиготными и ди-зиготными.
Он определили, что близнецы, рожденные
физические сходными, остаются похожими
и в дальнейшей жизни. Сходство не только
физическое, но и в уровне умственного
развития, личностных качествах, интересах.
Гальтон исследовал также и родословные
выдающихся людей, придя к выводу о наследственной
природе таланта.
Изучение родословных,
и близнецовый метод и сейчас
широко применяются для генетических
исследований человека. Эти методы
были успешно использованы и при
изучении поведенческих особенностей,
таких как стремление к новым впечатлениям,
склонность к агрессии, сексуальная ориентация
и многих других. В последние десятилетия
арсенал генетиков пополнился молекулярными
методами, позволившими не только выявить
влияние наследственности на те или иные
особенности поведения, но и найти гены,
которые определяют ряд психологических
характеристик человека. Изучение генома
– совокупности всех генов и межгенных
последовательностей ДНК – помогает приоткрыть
тайну человеческой индивидуальности.
Понять роль определенных генов в становлении
поведения человека помогают исследования
на животных – мышах, мухах дрозофилах
и даже совсем примитивных червях нематодах.
Однако генетические исследования поведения
обладают своими особенностями. Данные
одних исследователей зачастую не подтверждаются
другими исследователями – в отличие
от работ в других областях генетики, где
результаты гораздо менее противоречивы.
Это связано с тем, что многие формы поведения
зависят от большого числа генов и гораздо
больше подвержены влиянию внешней среды,
чем биохимические и морфологические
признаки. B 1923 году вышла в свет известная
работа И.П. Павлова «Новые исследования
по условным рефлексам», в которой автор
высказал мнение, будто приобретенные
в течение жизни условные рефлексы могут
передаваться по наследству потомкам,
превращаясь со временем во врожденные,
безусловные рефлексы. Другой русский
ученый, H.K. Кольцов, выступил с резкой
критикой представлений И.П. Павлова о
наследственной передаче благоприобретенных
признаков, в данном случае условных рефлексов.
H.K. Кольцов отметил, что И.П. Павлов никогда
не работал в области генетики и не представляет
себе всей сложности генетических проблем.
И.П.Павлов был вынужден согласиться с
критикой и отказался от своего предположения.
Это была, пожалуй, первая серьезная полемика
по вопросам наследования поведенческих
навыков, что можно понять, поскольку экспериментальные
исследования по генетике поведенческих
признаков, включая обучение, были начаты
лишь на заре XX века, при этом именно российские
ученые Кольцовской школы были в числе
первопроходцев.
Другие методы расчета наследуемости
Полнее всего, как показано выше, наследуемость после гибридизации рассчитывают по формулам Матера. Наследуемость в широком смысле можно также рассчитать только по F2, если допустить, что окружающая среда в одинаковой мере влияет и на поколение родителей, и на популяцию F2. Разница между средней ценностью варианс поколения родителей и F2 дает генотипическую вариансу. Наследуемость вычисляют по формуле:
Эту формулу применяют только для наследуемости в широком смысле, которая для показателя числа колосков в колосе составляет:

Если вместе с F2 и поколениями родителей в те же годы выращивают F1, то варианса F1 совместно с вариансами родительских поколений принимается за экологическую и вычитается из вариансы F2. Следует избегать использования вариансы F1, так как она часто обнаруживает сильный эффект супердоминантности и последовательного взаимодействия с окружающей средой, что почти всегда не находит своего отражения в F2.
Наследуемость можно рассчитывать и как регрессию селекционной ценности от фенотипической ценности:

Наследуемость также можно рассчитать и через коэффициент корреляции между селекционной и фенотипической ценностью:
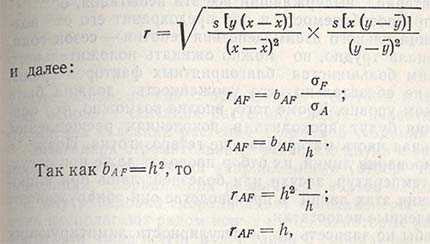
а это означает, что коэффициент корреляции между селекционной ценностью (А) и фенотипической ценностью (F) равен наследуемости. Следовательно:
Выведение формул показано в учебниках Фальконера, Матера и Джинкса и других авторов.
Учитывая, что наследуемость отдельных признаков имеет огромное значение для генетической ценности отбора, о ней еще будет сказано в главе о методах селекции.
Cookie файлы бывают различных типов:
Необходимые. Эти файлы нужны для обеспечения правильной работы сайта, использования его функций. Отключение использования таких файлов приведет к падению производительности сайта, невозможности использовать его компоненты и сервисы.








Файлы cookie, относящиеся к производительности, эффективности и аналитике. Данные файлы позволяют анализировать взаимодействие посетителей с сайтом, оптимизировать содержание сайта, измерять эффективность рекламных кампаний, предоставляя информацию о количестве посетителей сайта, времени его использования, возникающих ошибках.
Рекламные файлы cookie определяют, какие сайты Вы посещали и как часто, какие ссылки Вы выбирали, что позволяет показывать Вам рекламные объявления, которые заинтересуют именно Вас.
Электронная почта. Мы также можем использовать технологии, позволяющие отслеживать, открывали ли вы, прочитали или переадресовывали определенные сообщения, отправленные нами на вашу электронную почту. Это необходимо, чтобы сделать наши средства коммуникации более полезными для пользователя. Если вы не желаете, чтобы мы получали сведения об этом, вам нужно аннулировать подписку посредством ссылки «Отписаться» («Unsubscribe»), находящейся внизу соответствующей электронной рассылки.
Сторонние веб-сервисы. Иногда на данном сайте мы используем сторонние веб-сервисы. Например, для отображения тех или иных элементов (изображения, видео, презентации и т. п.), организации опросов и т. п. Как и в случае с кнопками доступа к социальным сетям, мы не можем препятствовать сбору этими сайтами или внешними доменами информации о том, как вы используете содержание сайта.
Наследуемость психологического признака
Люди отличаются
друг от друга рядом психологических
характеристик. Эти различия
вызваны как неодинаковыми условиями
жизни, так и несходными генотипами, поскольку
генотипы людей содержат разные формы
генов. Соотносительный вклад наследственности
и среды в разнообразие людей по психологическим
свойствам и поведению изучает психогенетика.
Для оценки влияния наследственности
и среды на поведение человека ученые
сравнивают людей, имеющих различную степень
генетической общности (однояйцевых и
многояйцевых близнецов, родных и сводных
братьев и сестер, детей и их биологических
и приемных родителей).
Многие гены
существуют в нескольких формах, подобно
тому, как есть разные формы гена,
определяющего цвет глаз. Некоторые
гены имеют десятки форм. Генотип
конкретного человека содержит два
экземпляра каждого гена, формы которых могут быть разными, а могут
быть и одинаковыми. Один унаследован
от отца, другой — от матери. Сочетание
форм всех генов уникально для каждого
человеческого организма. Эта уникальность
лежит в основе генетически обусловленных
различий между людьми. Вклад генетических
различий в разнообразие людей по психологическим
свойствам отражает показатель, называемый
«коэффициент наследуемости». Например,
для интеллекта коэффициент наследуемости
составляет, по меньшей мере, 50%. Это не
означает, что 50% интеллекта дано человеку
от природы, а остальные 50% нужно добавить
путем обучения, тогда интеллект будет
100 баллов. Коэффициент наследуемости
не имеет отношения к конкретному человеку.
Его вычисляют, чтобы понять, в чем причина
отличия людей друг от друга: возникают
ли различия из-за того, что у людей неодинаковые
генотипы, или потому, что их по-разному
учили. Если бы коэффициент наследуемости
интеллекта оказался близок к 0%, то можно
было бы сделать вывод, что только обучение
формирует различия между людьми, и применение
одних и тех же воспитательных и образовательных
приемов к разным детям будет всегда приводить
к одним и тем же результатам. Высокие
значения коэффициента наследуемости
означают, что даже при одинаковом воспитании
дети будут отличаться друг от друга в
силу своих наследственных особенностей.
Конечный результат, однако, не предопределен
генами. Известно, что дети, усыновленные
в благополучные семьи, по уровню интеллектуального
развития оказываются близки к своим приемным
родителям и могут значительно превосходить
биологических. В чем же тогда выражается
влияние генов? Поясним это на примере
конкретного исследования.*
Ученые обследовали
две группы приемных детей. Условия
в приемных семьях были у всех одинаково
хорошими, а биологические матери
детей различались по уровню интеллекта.
Биологические матери детей из первой
группы имели интеллект выше среднего.
Примерно половина детей из этой группы
продемонстрировала интеллектуальные
способности выше средних, другая половина
— средние. Биологические матери детей
второй группы имели несколько сниженный
(но в пределах нормы) интеллект. Из этой
группы 15% детей имели такие же невысокие
оценки интеллекта, у остальных детей
уровень интеллектуального развития соответствовал
среднему. Таким образом, при одинаковых
условиях воспитания в приемных семьях,
интеллект детей, в определенной степени,
зависел от интеллекта их кровных матерей.
Приведенный пример
может служить иллюстрацией существенных
различий между понятием наследуемости
психологических качеств и наследуемости некоторых физических особенностей
человека, таких как цвет глаз, кожи и т.д.
Даже при высоком уровне наследуемости
психологического признака генотип не
предопределяет его конечного значения.
От генотипа зависит, как ребенок будет
развиваться в определенных условиях
среды. В некоторых случаях генотип задает
«пределы» выраженности признака.
Источники
Cullis B.R., Thomson F.M., Fisher J.A., Gilmour A.R., Thompson R. The analysis of the NSW wheat variety database. I. Modelling trial error variance // Theoretical and Applied Genetics. 1996. No. 92. P. 21–27. doi:10.1007/BF00222947
Cullis B.R., Smith A.B., Coombes N.E. On the design of early generation variety trials with correlated data // Journal of agricultural, biological, and environmental statistics. 2006. No. 11 (4). P. 381.
Falconer D.S., Mackay T.F.C. Introduction to quantitative genetics. 4th ed. Pearson Prentice Hall. NJ.: Upper Saddle River, 2005.
Holland J.B., Nyquist W.E., Cervantes-Martínez C.T. Estimating and interpreting heritability for plant breeding: An update // Plant Breeding Reviews. 2003. P. 9–112. doi:10.1002/9780470650202.ch2
Knight R.L. Dictionary of genetics, including terms used in cytology, animal breeding and evolution. 1948.
Kor Oldenbroek and Liesbeth van der Waaij, 2015. Textbook Animal Breeding and Genetics for BSc students. Centre for Genetic Resources The Netherlands and Animal Breeding and Genomics Centre, 2015. Groen Kennisnet. URL: https://wiki.groenkennisnet.nl/display/TAB/
Lourenço V.M. et al. A robust DF-REML framework for variance components estimation in genetic studies // Bioinformatics. 2017. No. 33 (22). P. 3584–3594.
Oakey H., Verbyla A., Pitchford W., Cullis B., Kuchel H. Joint modeling of additive and non-additive genetic line effects in single field trials // Theoretical and Applied Genetics. 2006. No. 113. P. 809–819. doi:10.1007/ s00122-006-0333-z
Piepho H.-P., Möhring J. Computing heritability and selection response from unbalanced plant breeding trials // Genetics. 2007. No. 177. P. 1881–1888. doi:10.1534/genetics.107.074229
Schmidt P. et al. Estimating Broad‐Sense Heritability with Unbalanced Data from Agricultural Cultivar Trials // Crop Science. 2019. No. 59 (2). P. 525–536.
Schmidt P. et al. Heritability in plant breeding on a genotype-difference basis // Genetics. 2019. No. 212 (4). P. 991–1008.
Smith A., Cullis B., Gilmour A. The analysis of crop variety evaluation data in Australia // Australian and New Zealand journal of statistics. 2001. No. 43. P. 129–145. doi:10.1111/1467-842X.00163
Walsh B., Lynch M. Evolution and selection of quantitative traits. 1st ed. Oxford, UK: Oxford Univ. Press, 2018. doi:10.1093/ oso/9780198830870.001.0001









Наследуемость
Отбор проводится на основе фенотипической ценности, поэтому важно знать, с какой вероятностью отобранные фенотипы дадут тождественное потомство. Если значение генетической вариансы по определенному признаку велико, а значение экологической вариансы мало, можно ожидать, что потомство в немалой мере будет таким же, как и отобранные фенотипы
И наоборот, если генетическая варианса мала, а экологическая велика, потомство по ценности может значительно отличаться от отобранных фенотипов.
Сходство между родителями и их потомством в огромной степени зависит от компонентов генетической вариансы (VA + VD). Если речь идет об аддитивном компоненте генетической вариансы (VA), то здесь фенотипы родителей представляют собой надежные индикаторные признаки своих генотипов и, следовательно, дадут сходное потомство. При доминантном компоненте генетическая варианса (VD) будет давать отличное от родительских фенотипов потомство, и это зависит от характера межаллельного взаимодействия.
Соотношение между генотипической вариансой и суммарной фенотипической вариансой называется наследуемостью (Н, или h2) какого-то признака определенной популяции и обозначается:
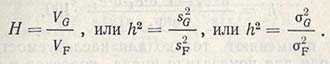
Это наследуемость в широком смысле. Наследуемость в узком смысле — соотношение только между аддитивным компонентом генотипической вариансы и суммарной фенотипической вариансой:
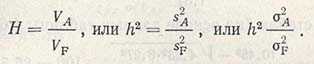
Наследуемость по признаку числа колосков в колосе в проанализированном примере составляет:
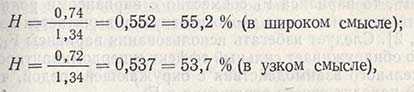
т.е. показатель наследуемости относительно высок. Следовательно, генетические различия между родителями были велики, и в последующих поколениях с помощью отбора можно будет выделить генотипы с большим числом колосков (из сорта Безостая 1) и объединять их с генотипами, имеющими большое число зерен (из Siete Cerros). Но так как величина экологической изменчивости весьма значительна, то этого может быть достаточно, чтобы истинная ценность генотипов оказалась скрытой и произошел отбор модификаций, которые в следующем поколении не дадут растений с большим числом колосков.
Как уже было сказано, для успешного проведения отбора наибольшее значение имеет аддитивный компонент генетической изменчивости, названный поэтому селекционной ценностью. Фальконер считает, что наследуемость выражается в пригодности фенотипической ценности служить ориентиром селекционной ценности или же она отражает степень совпадения между фенотипической и селекционной ценностью.
Происхождение использования этого понятия
Это понятие было введено в публичные дебаты в 1969 году Артуром Дженсеном в статье, озаглавленной « Насколько мы можем повысить IQ и учебные достижения?» » Опубликовано в Harvard Educational Review . В этой знаменитой статье Дженсен утверждает, что программы компенсационного образования в Соединенных Штатах, направленные на повышение успеваемости детей из неблагополучных семей (особенно из бедных семей, принадлежащих к этническим меньшинствам), не увенчались успехом. Дают эффект, потому что они основаны на неправильных предпосылках . Опираясь, в частности, на работу Сирила Берта ( « Самая удовлетворительная попытка оценить отдельные компоненты дисперсии — это работа сэра Сирила Берта (1955, 1958) » ), а также на обзор, опубликованный Эрленмейер-Кимлинг и Джарвик в 1963 году. На основании исследований семей и близнецов, направленных на оценку наследуемости интеллектуальных способностей, Дженсен приходит к выводу, что «наследуемость интеллекта» составляет в среднем около 80%.
Объединив эту оценку с другими данными, он затем утверждает, что очевидно, что различия в IQ, наблюдаемые между социально-экономическими классами, а также между расами в Соединенных Штатах, по крайней мере частично вызваны генетическими различиями между этими группами. В отношении расовых различий он рассуждает следующим образом:
Эта статья, а также последующее использование оценок наследуемости, основанных на том же типе рассуждений, например, сделанных в The Bell Curve в 1994 году, а также совсем недавно в книге журналиста Николаса Уэйда « Трудное наследование: гены, раса и история человечества » в 2014 г. вызвал резкую критику в научном сообществе. В дополнение к сомнению в достоверности использованных оценок наследуемости IQ было указано, что эта информация не позволяет сделать какой-либо вывод о причинах наблюдаемых различий между двумя разными популяциями, живущими в разных средах.
Оценка наследуемости IQ
Наследуемость в IQ баллов или других показателей интеллекта (г фактора, успеваемость и т.д.) является предметом многочисленных оценок в различных популяциях и с различными методами.
Оценка классическими методами изучения родственников и близнецов
Классические исследования родственников или близнецов часто давали оценки наследуемости IQ во взрослом возрасте порядка от 50 до 80%. Однако в этих методах оценки накапливаются систематические ошибки, которые приводят к переоценке наследуемости, включая исследования близнецов, которые иногда ошибочно представляются надежными.
Оценка с помощью моделей, учитывающих эффекты большого количества SNP.
С 2000-х годов были разработаны методы оценки наследуемости, основанные на геномных данных по большим выборкам особей. Общий принцип метода , который до сих пор наиболее часто используемый состоит в построении математической модели статистического эффекта генотипа как сочетание статистических эффектов более или менее большого подмножества моно- полиморфизмов. Нуклеотиды ( SNP).
Только учет нескольких десятков тысяч SNP позволяет воссоздать с помощью специальной статистической модели существенную часть дисперсии, приписываемой генотипам в выборке используемой популяции, то есть так сказать называется геномной наследуемостью, иногда приближающейся к 50%. Однако там, где это уместно, эти модели страдают от переобучения , то есть от переобучения модели данным. В этом случае модель «объясняет» большую часть дисперсии по SNP, но только потому, что она включала случайный шум, присутствующий в данных выборки, которая использовалась для ее построения (поэтому часть этой предполагаемой наследуемости является фиктивной) . Для ограничения переобучения рекомендуется выполнять перекрестную проверку модели на других образцах. Реализация этих перекрестных проверок в исследованиях на людях остается недостаточной, и на самом деле часть дисперсии, объясняемая этими моделями, часто резко падает, когда они применяются к другой выборке.
Так, в Hill et al. Например, в 2018 году авторам удалось построить математическую модель, объясняющую 25,44% дисперсии показателя интеллекта в исходной выборке, но когда эта модель была применена к трем выборкам репликации, только 3,64% — 6,84% дисперсия показателей интеллекта была предсказана моделью.
Аналогичным образом модель, построенная Ли и др. 2018, основанный на выборке из 1,1 миллиона человек европейского генетического происхождения с использованием около 250 000 SNP, уловил только 12,7% дисперсии по количеству лет исследований и 6,9% дисперсии когнитивных функций в одной когорте репликации, а также 10,6 % и 9,7% соответственно в другом. Кроме того, по словам авторов, модель потеряла 85% своей предсказательной силы при применении к выборке афроамериканцев. Авторы далее подчеркивают, что специфический эффект генетических вариантов, вероятно, переоценивается моделью из-за «корреляции между уровнем образования и образовательной средой, способствующей академическому успеху» . В 2019 году доля дисперсии, предсказанная этой моделью, была самой высокой из когда-либо достигнутых с помощью этого типа подхода с учетом всех когнитивных или поведенческих характеристик вместе взятых.







Оценка методом RDR ( неравновесная регрессия родства )
Метод RDR — это недавний метод, также основанный на геномных данных, направленный на компенсацию определенных недостатков методов, основанных на моделировании статистических эффектов SNP. Он оценивает наследственность, исследуя, как фенотипическое сходство между людьми изменяется в зависимости от их генетической близости, измеряемой долей генома, разделяемой двумя людьми, поскольку они унаследованы от общего предка. Он был проведен впервые в 2018 году на выборке из почти 55000 исландцев, и в результате оценки наследуемости были значительно ниже, чем оценки, полученные с помощью существующих методов. В частности, наследуемость уровня исследований была оценена в этой выборке в 17% против соответственно 52%, 29% и 40% для трех других методов и 43% для исследования референтных близнецов.
Заключение и рекомендации
настоящем руководстве представлена правильная и корректная интерпретация понятия наследуемости, а также рассмотрены распространенные заблуждения, которых следует избегать. Кроме того, проведен обзор основных методов расчета наследуемости, анализированы их преимущества и недостатки. В частности, метод Куллиса был рекомендован как надежный для учета несбалансированных наборов данных. Также были рассмотрены методы Пьефо и Уолша и Линча , которые также считаются надежными, однако требуют дополнительных вычислительных усилий.
Руководство предоставляет полезную информацию для исследователей и селекционеров, помогая им понять и правильно интерпретировать показатели наследуемости
Оно подчеркивает важность выбора надежного метода расчета, особенно при работе с несбалансированными данными. Внимательное изучение обзора методов позволит исследователям определить наиболее подходящий и эффективный способ оценки наследуемости в своих исследованиях