

ТИПЫ АНАТОМИЧЕСКОГО СТРОЕНИЯ СТЕБЛЯ
В зависимости от того, какие меристемы участвуют в образовании стебля, различают первичное и вторичное строение стебля. Строение стебля, сформировавшееся в результате деятельности первичных меристем (апикальная меристема, перицикл, прокамбий), называется первичным. Стебли однодольных растений имеют первичное строение на всем протяжении жизни. Стебли двудольных и голосеменных растений имеют первичное строение лишь на ранних этапах развития. За счет апикальной меристемы формируется покровная ткань – эпидерма и первичная кора, за счет перицикла и прокамбия – центральный цилиндр (в частности прокамбий, образует первичные элементы флоэмы и ксилемы).
На определенных этапах развития у двудольных и голосеменных растений первичное строение сменяется на вторичное. Переход к вторичному строению связан с заложением камбия и феллогена. Камбий возникает в центральном цилиндре из клеток прокамбий и паренхимы. В свою очередь камбий образуют вторичные элементы флоэмы и ксилемы. Особенно много образуется ксилемы, за счет чего стебель значительно утолщается. Феллоген закладывается под эпидермой. За счет его деятельности формируется вторичная покровная ткань – пробка. После образования пробки эпидерма отмирает и слущивается.
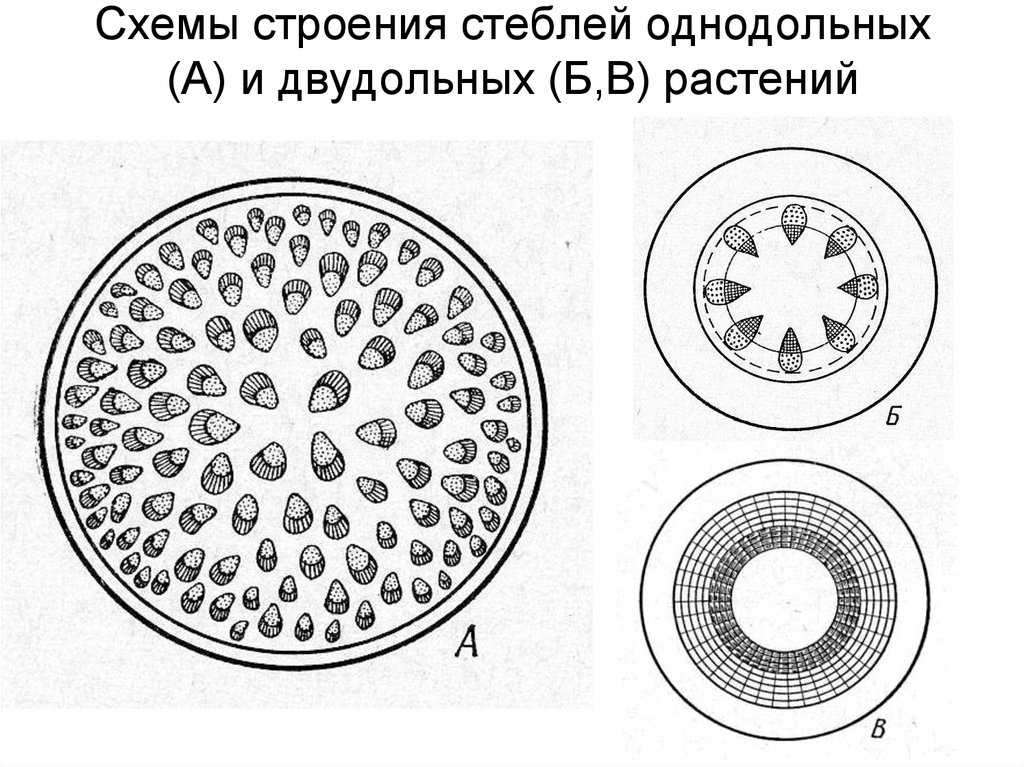
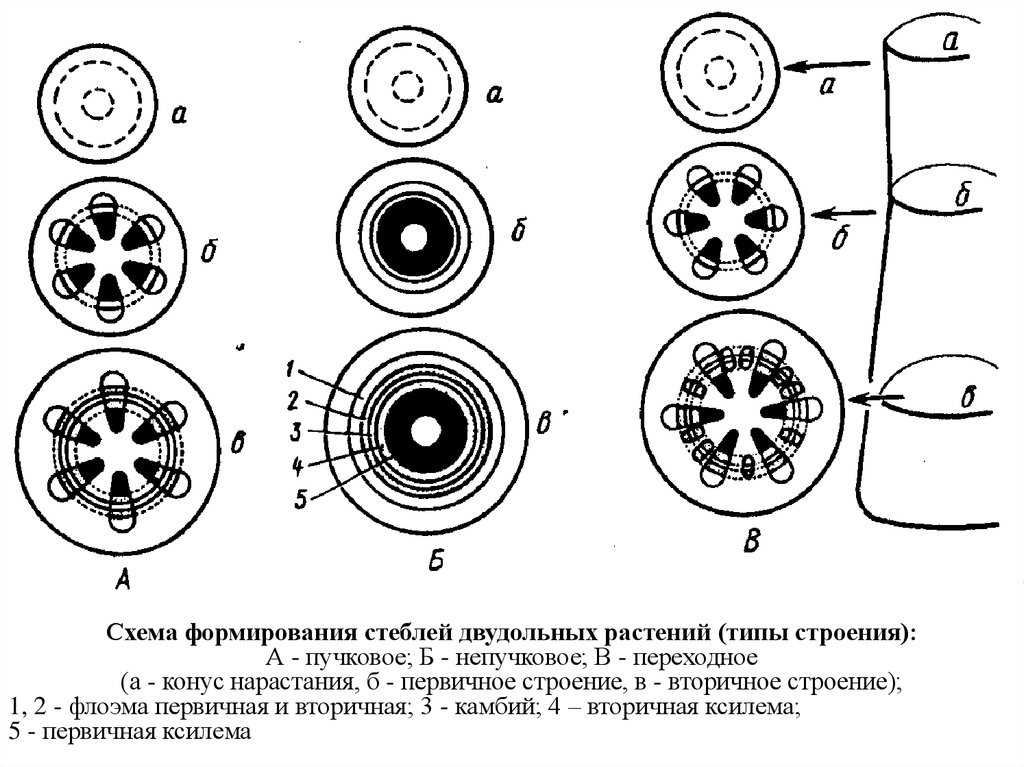
В зависимости от способов заложения прокамбия и камбия различают пучковое, непучковое и переходное строение стебля (рисунок 6).
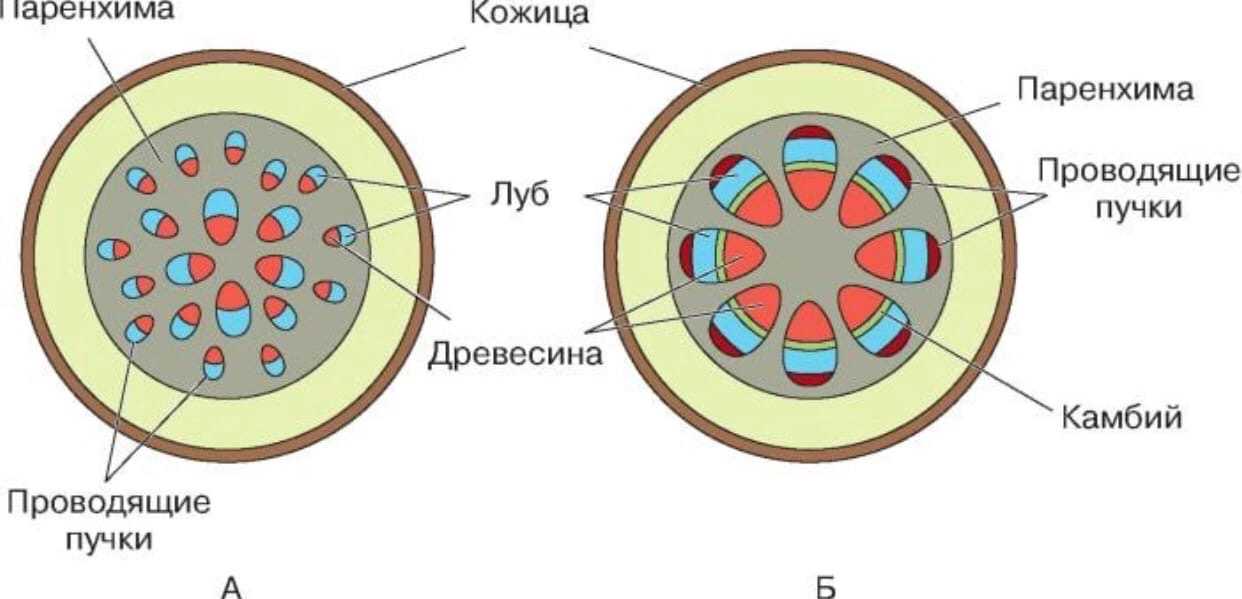
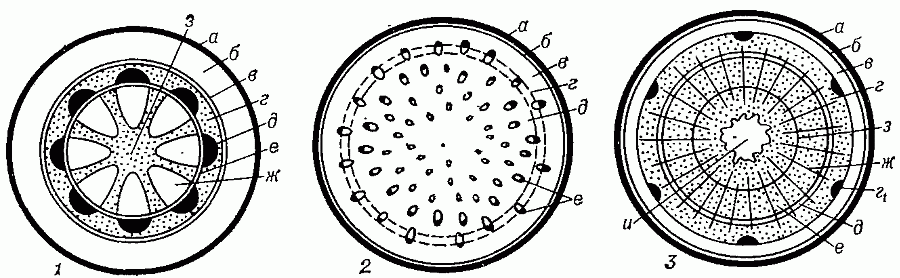
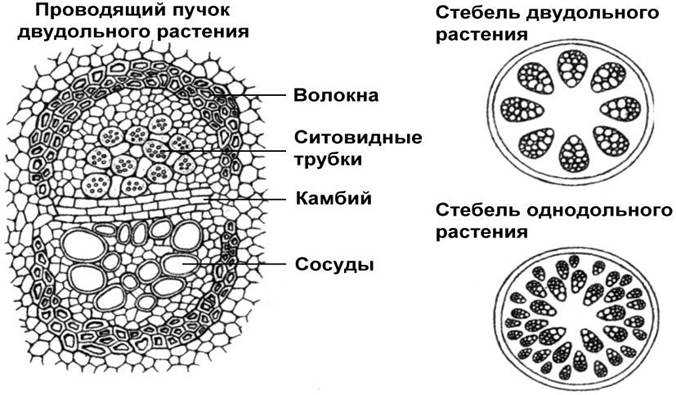
Пучковоестроение стебля встречается у некоторых травянистых растений. Пучковое строение формируется в том случае, если прокамбий закладывается отдельными тяжами. Заложенные в конусе нарастания прокамбиальные тяжи располагаются в один круг по периферии центрального цилиндра. Каждый Прокамбиальный тяж превращается в коллатеральный пучок, состоящий из первичной ксилемы и первичной флоэмы. У однодольных растений прокамбиальные тяжи полностью превращаются в элементы флоэмы и ксилемы. Таким образом, проводящие пучки у однодольных растений будут закрытыми коллатеральными (например, у кукурузы и ржи). У двудольных растений остается полоска прокамбия, которая превращается в полоску пучкового камбия. Клетки пучкового камбия, делясь, дают новые (вторичные) элементы ксилемы и флоэмы. Проводящие пучки в этом случае будут открытые коллатеральные (например, у клевера, кирказона). Кроме того, деятельность пучкового камбия стимулирует паренхиму, разделяющую пучки, которая начинает делиться, давая межпучковый камбий. Межпучковый камбий в свою очередь производит паренхиму сердцевинных лучей или склеренхиму.
Пучковое строение стебля может с возрастом смениться непучковым, сплошным. Таким переходнымстроением отличаются стебли подсолнечника, георгины, клещевины, петрушки и др. Вначале стебель формируется так же, как у клевера, но межпучковый камбий откладывает не паренхиму или склеренхиму, а ксилему и флоэму. Формируются добавочные проводящие пучки. Постепенно все пучки могут слиться в сплошные цилиндры флоэмы и ксилемы (цилиндр ксилемы окружает цилиндр флоэмы, между ними камбий). В верхней (молодой) части стебель имеет пучковое строение, у основания (в старой части) — непучковое.
Непучковое строение свойственно стеблям древесных растений (например, береза, липа, сосна) и некоторым травам (например, лен). В конусе нарастания этих растений прокамбиальные тяжи настолько сближены, что образуют почти сплошной цилиндр (на поперечном срезе он имеет вид кольца). Прокамбиальный цилиндр, дифференцируясь, дает к центру цилиндр первичной ксилемы, к периферии — цилиндр первичной флоэмы. Затем на месте прокамбия закладывается камбий, также в виде цилиндра. Вторичные изменения связаны с его работой. Внутрь камбий формирует цилиндр вторичной ксилемы (древесины), наружу откладывает цилиндр вторичной флоэмы (луб). Местами камбий откладывает в обе стороны паренхимные клетки сердцевинных лучей.
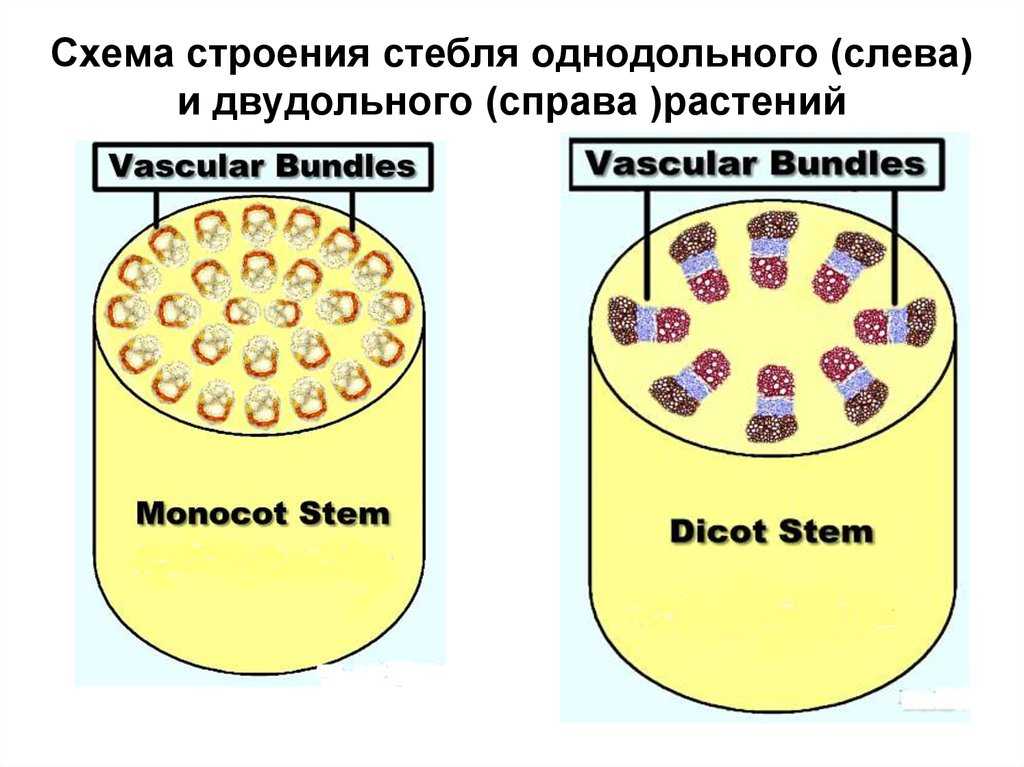
Таким образом, у однодольных растений анатомическое строение стебля – первичное пучковое, пучки закрытые коллатеральные. У двудольных и голосеменных растений вторичное: у травянистых растений все три типа – пучковое (пучки открытые коллатеральные), непучковое и переходное; у древесных растений всегда непучковое строение.
Рисунок 6. А – пучковое строение, Б – переходное строение, В – непучковое строение; I – заложение прокамбия, II – первичное строение III – заложение камбия и вторичное строение; 1- прокамбий, 2- первичная флоэма, 3 – первичная ксилема, 4 – вторичная флоэма, 5 – вторичная ксилема, 6 – пучковый камбий, 7 – межпучковый камбий.
Анатомия стебля
Основные функции стебля — проведение воды и питательных веществ. Дополнительными функциями могут быть фотосинтез, обычно происходящий в молодых стеблях растений, и отложение запасных питательных веществ в многолетних стеблях.
Анатомическое строение стебля растений обусловлено его главными функциями.
Для него характерно развитие механической и проводящей тканей растений. Кроме того, для стебля характерна сложная система меристем — верхушечные, боковые и вставочные, определяющие нарастание его в течение длительного времени и возникновение новых органов.
Стебель растения возникает из апикальной меристемы, из которой дифференцируются три слоя тканей: покровная, проводящая, основная.
Сначала эти ткани представлены так называемой первичной меристемой (промеристемой), состоящей из протодермы, прокамбия и основной меристемы.
Первичные проводящие ткани развиваются из прокамбия.
Первые элементы флоэмы дифференцируются из наружных, расположенных к периферии клеток прокамбия. Первичная флоэма представлена тонкостенными недолговечными удлиненными клетками и носит название протофлоэмы, а наружные клетки ее могут быть представлены механическими волокнами.
Первичные элементы ксилемы — трахеиды, реже сосуды с кольчатыми и спиральными утолщениями стенок — возникают позже из внутренних клеток прокамбия и определяются в целом как протоксилема.
В ее состав помимо проводящих элементов входят паренхимные клетки.
Позже внутрь от протофлоэмы дифференцируется имеющая более или менее типичное для флоэмы строение. Наружу от протоксилемы формируется метаксилема, состоящая из трахеид или трахей с более утолщенными одревесневшими стенками.
Таким образом, за счет деятельности прокамбия и остальной меристемы апекса возникает первичное строение стебля растения.
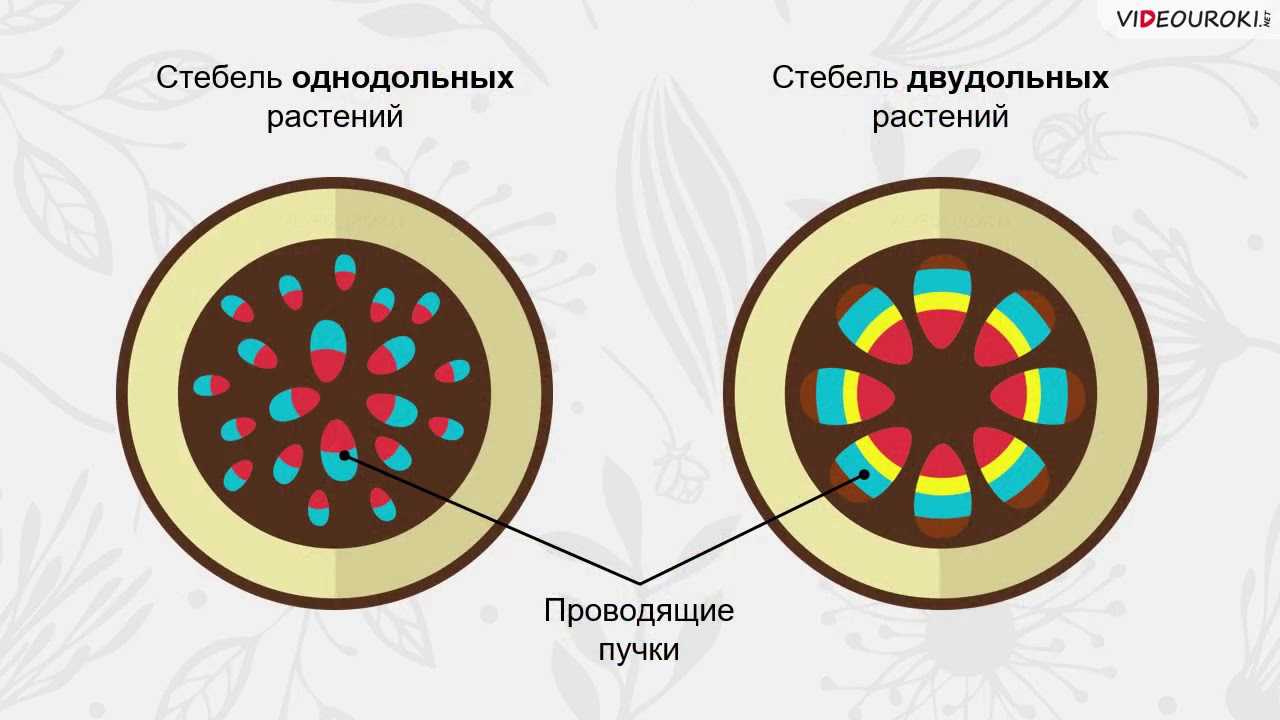
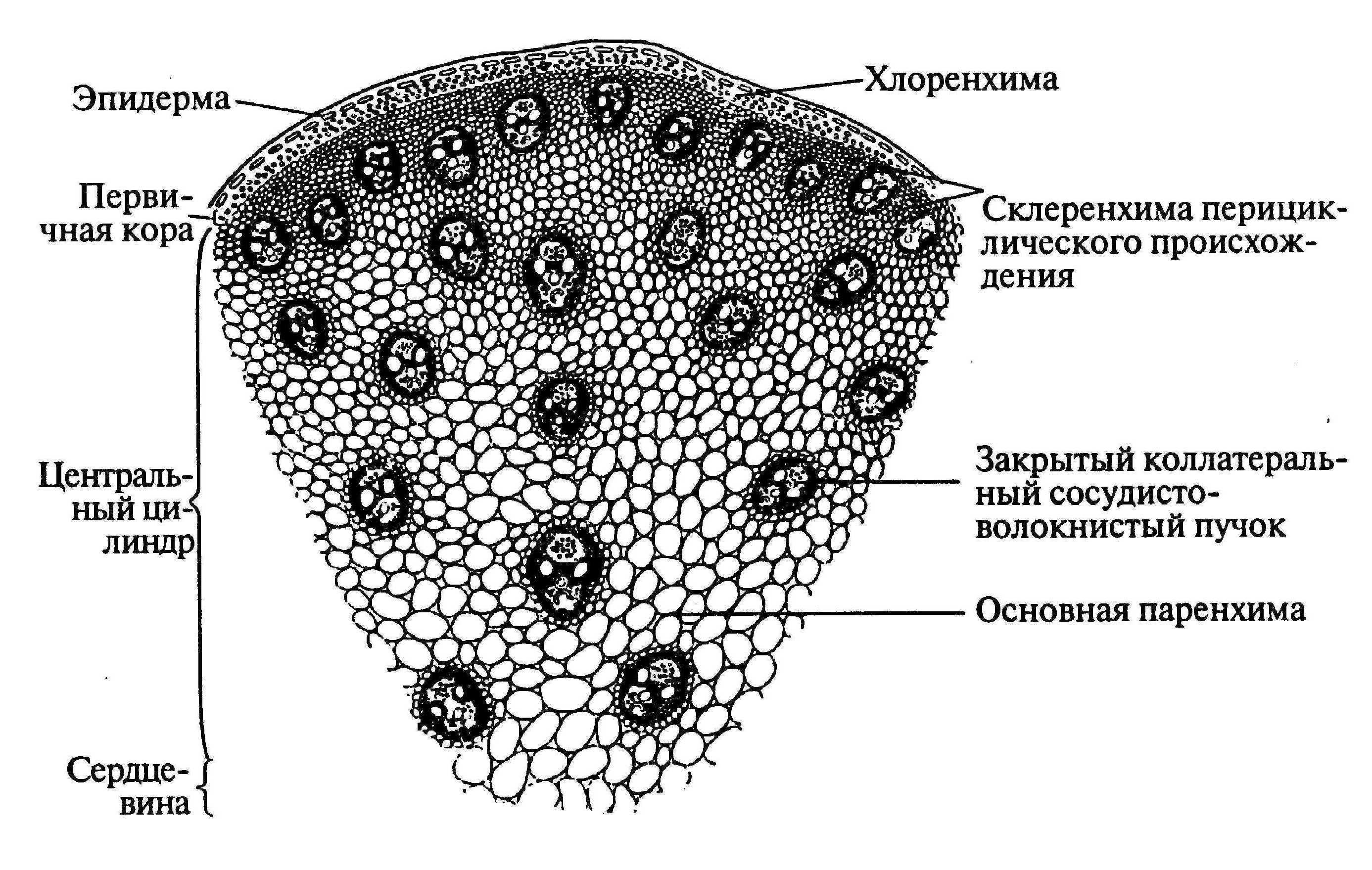
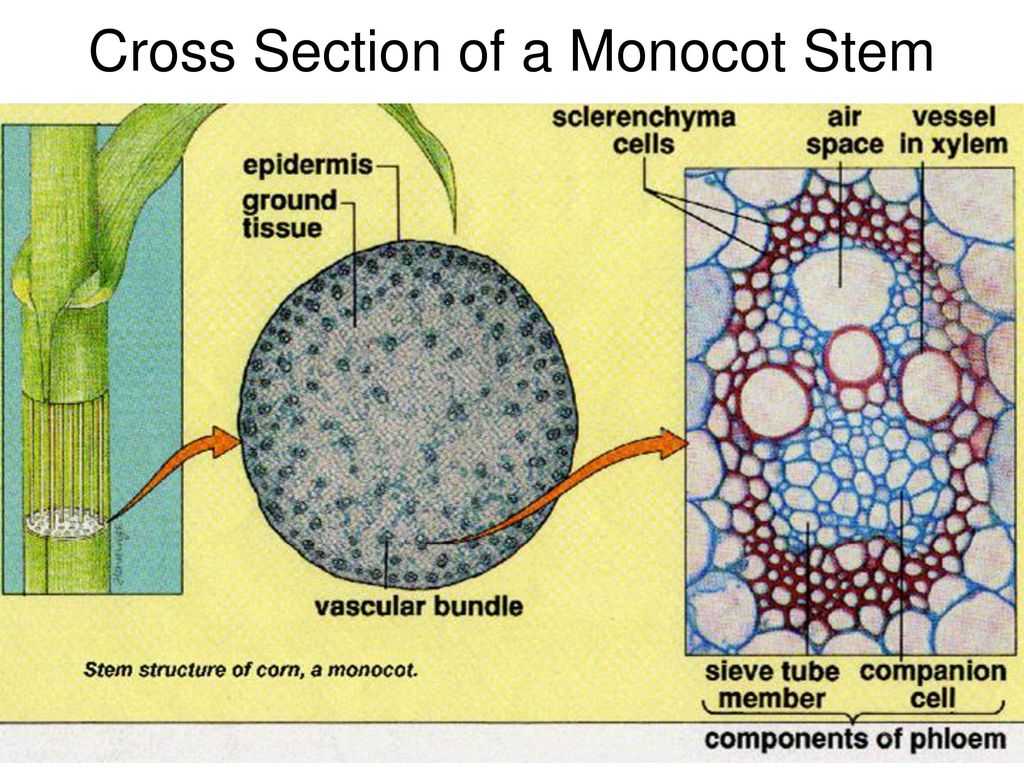
У однодольных растений весь прокамбий дифференцируется в элементы первичных проводящих тканей.
Первичное анатомическое строение стебля
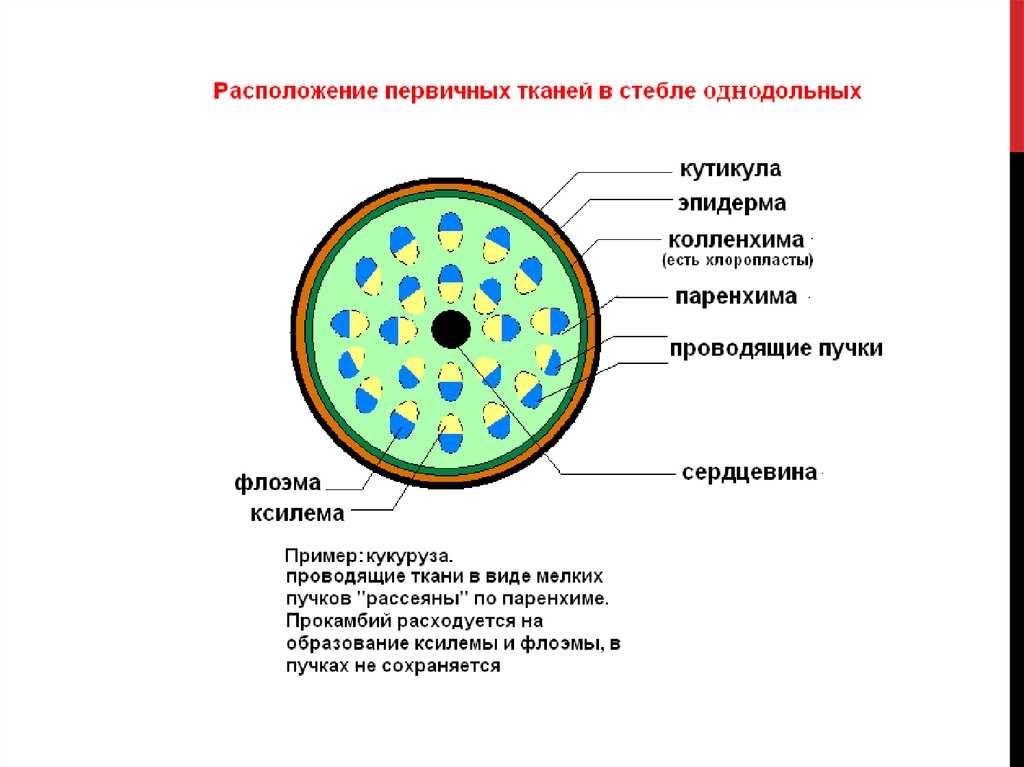
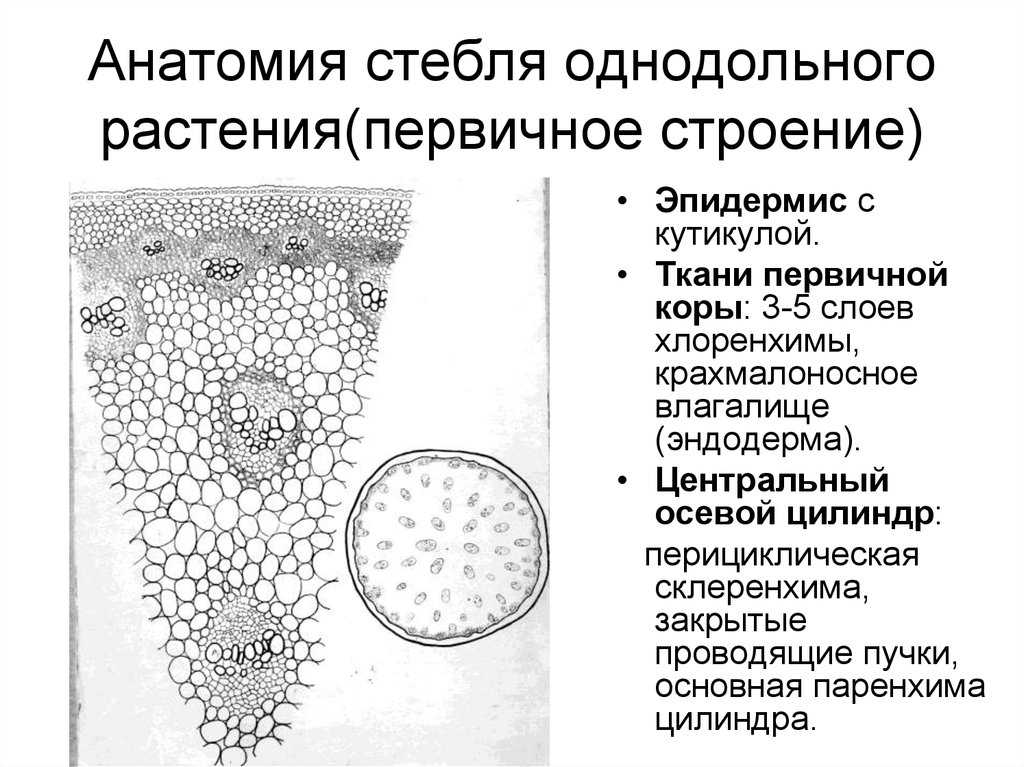
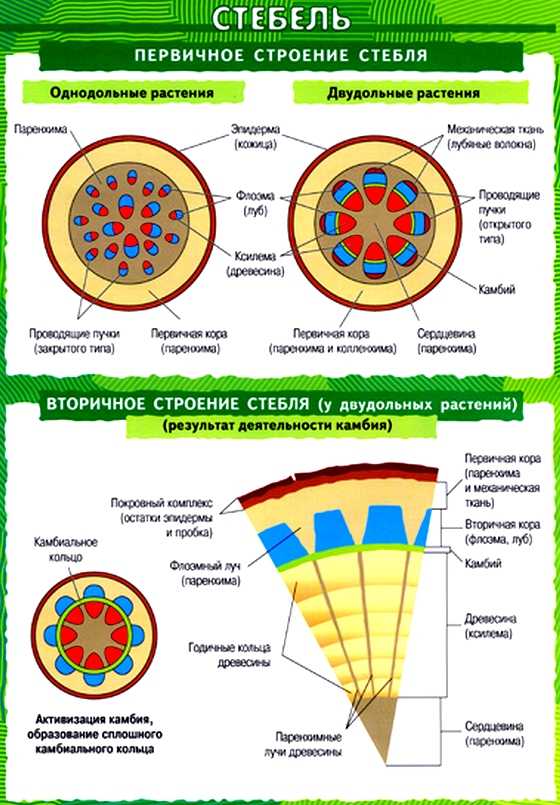
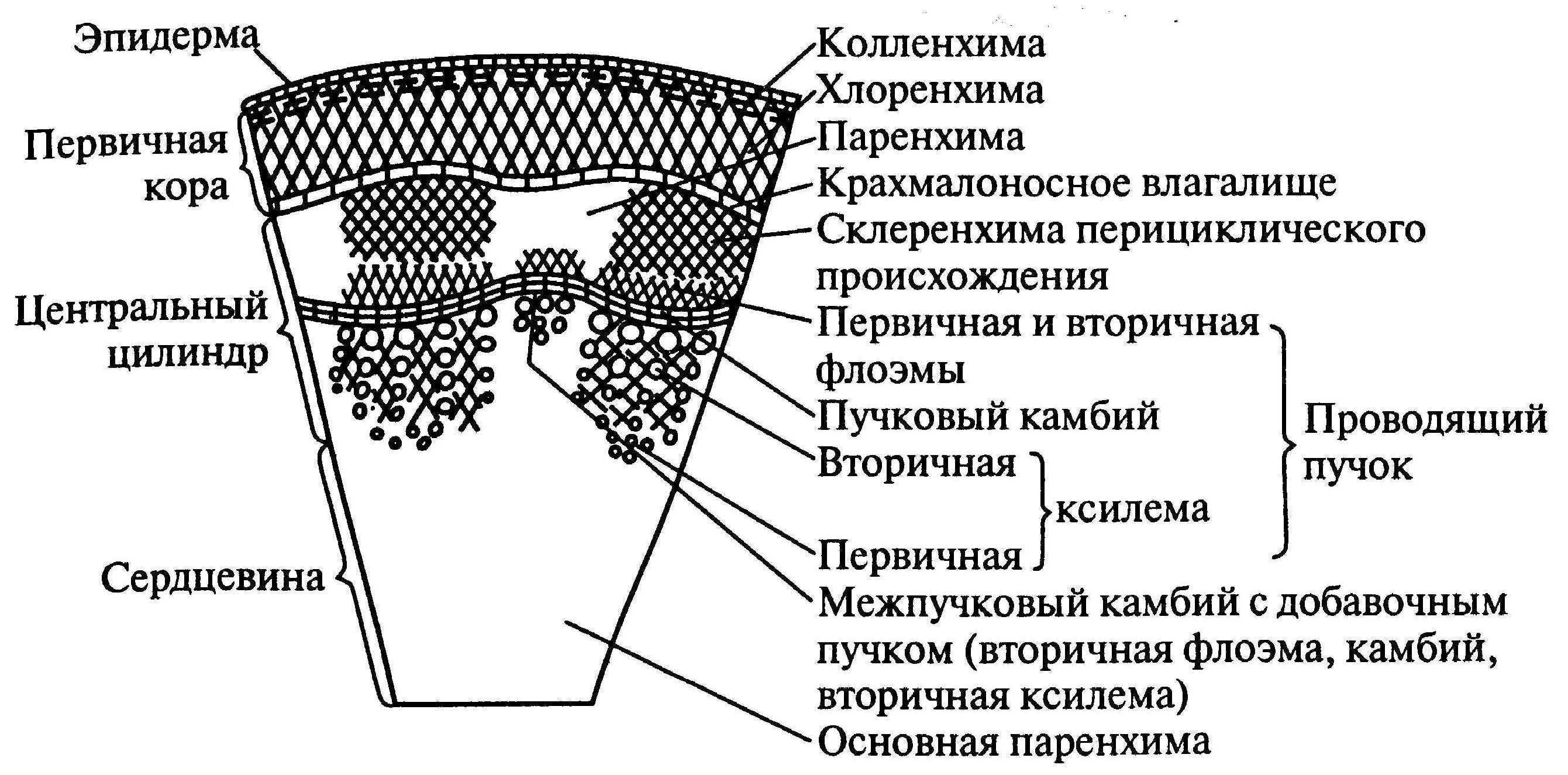
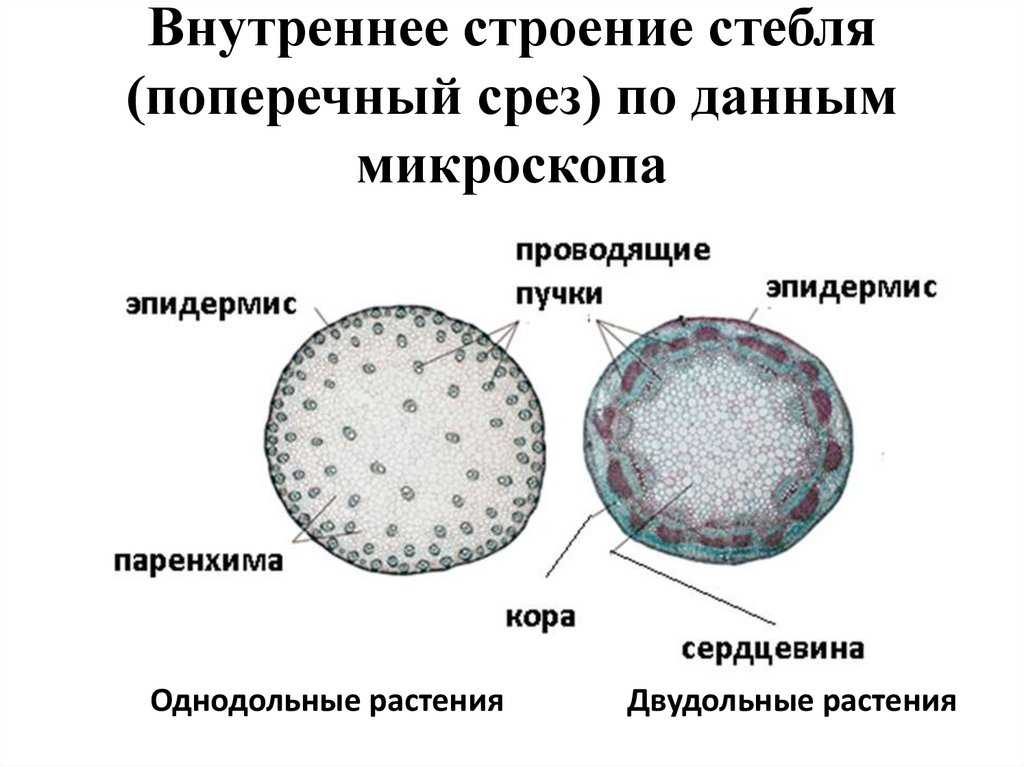
В стебле первичного строения выделяют следующие анатомо-топо- графические зоны: покровную ткань, первичную кору, центральный цилиндр и сердцевину (рис. 87).
Покровная ткань представлена эпидермой, защищающей стебель от потери воды. В располагающейся под эпидермой первичной коре хорошо выражена фотосинтезирующая паренхима (хлоренхима) и колленхима (у двудольных растений). Экзодерма и мезодерма в отличие от эндодермы обособлены слабо. Эндодерма (внутренний слой клеток первичной коры) содержит крахмальные зерна (амилопласты). Поэтому ее часто называют крахмалоносным влагалищем. Так как крахмальные зерна могут оседать под действием силы тяжести, существует гипотеза, что эндодерма может играть роль в геотропической реакции побега, являясь своеобразным «органом равновесия». Содержащийся в эндодерме крахмал растение не использует даже при голодании.
Как и в корне, центральный цилиндр стебля начинается с перицикла. Но в отличие от корня, где клетки перицикла долго сохраняют меристематическую активность, в стебле перицикл представлен уже образованными им постоянными тканями — склеренхимой и паренхимой.
Рис. 87. Первичное строение стебля (по Л.С. Родман, 2001 с изм.):
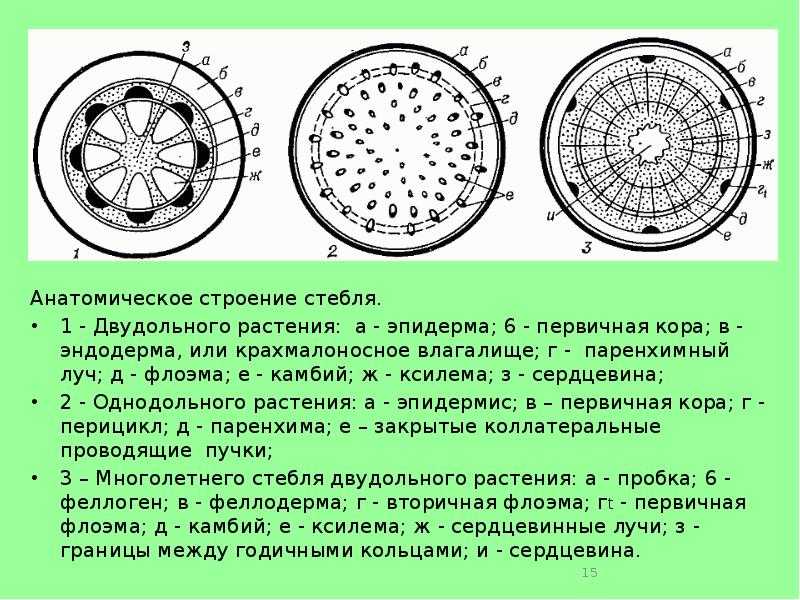
I — первичная кора;
II — центральный цилиндр;
III — сердцевина: 1 — эпидерма;
- 2 — колленхима; 3 — хлоренхима; 4 — крахмалоносное влагалище (эндодерма); 5— склеренхима;
- 6 — первичная флоэма;
- 7— закладывающийся камбий;
- 8 — первичная ксилема;
- 9 — паренхима сердцевины
Склеренхимные волокна образуют сплошной слой или располагаются рядом с проводящими пучками кнаружи от них.
Основу центрального цилиндра составляют проводящие пучки, сформировавшиеся из тяжей прокамбия и располагающиеся в паренхиме центрального цилиндра.
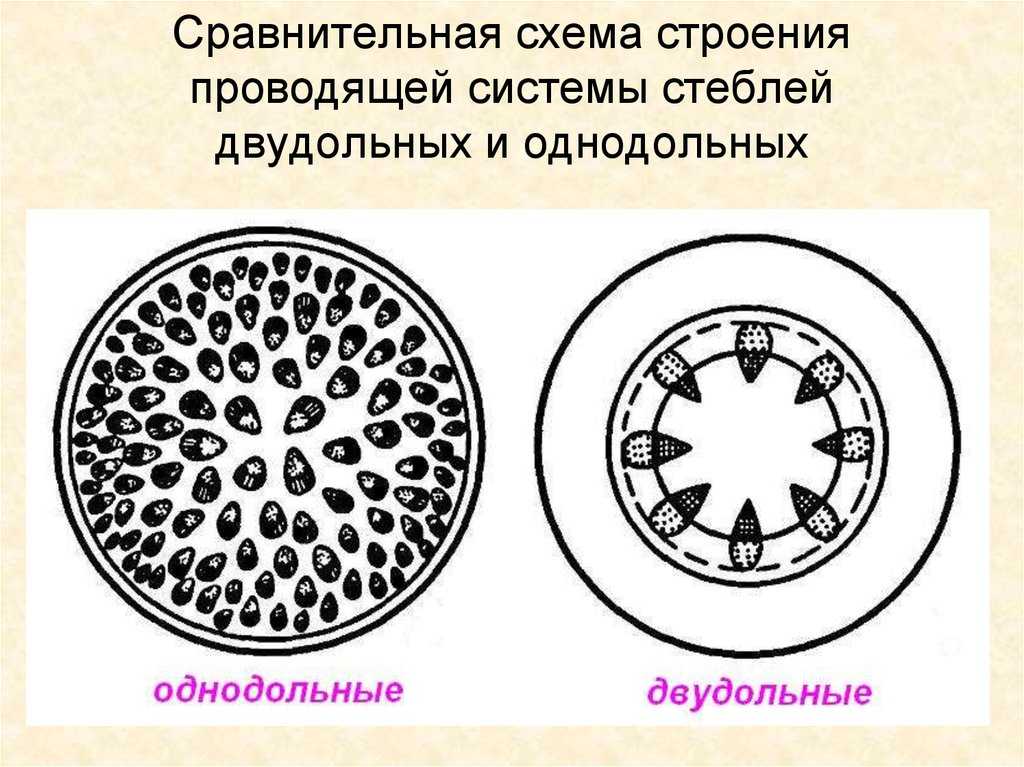
При образовании пучка дифференциация клеток прокамбия идет центростремительно: первичная ксилема развивается от внутренней части пучка к центру, первичная флоэма — навстречу ей. Если прокамбий полностью превращается во флоэму и ксилему, образуется закрытый коллатеральный пучок. Такие пучки типичны для однодольных растений. Если между флоэмой и ксилемой остается слой прокамбия, то он может дать начало камбию, который начнет формировать вторичные проводящие комплексы, и пучок будет открытым. При заложении прокамбия отдельными тяжами формируется пучковое строение стебля.
Если прокамбий закладывается в апексе побега в виде полого тонкостенного цилиндра, то развивающиеся из него первичные флоэма и ксилема тоже представлены вложенными друг в друга цилиндрами. Такое строение стебля называют непучковым, или сплошным (хвойные, двудольные покрытосеменные). При любом типе строения в стебле раньше формируются элементы флоэмы.
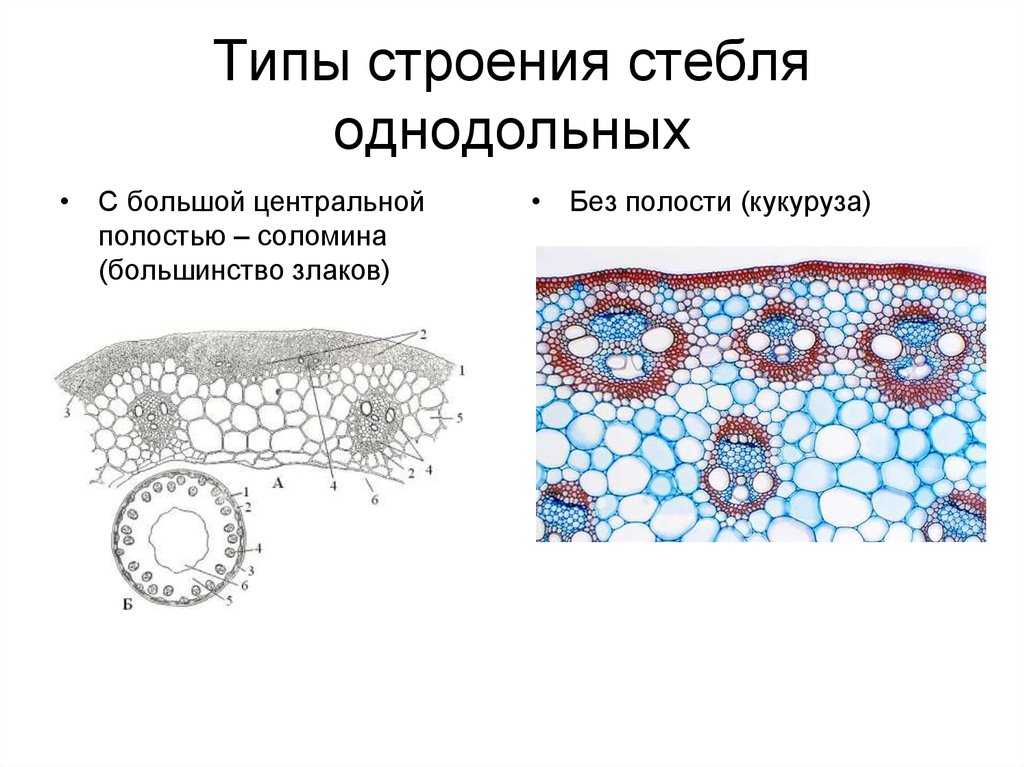
В центре стебля в отличие от корня всегда находится лучше или хуже развитая сердцевина. Довольно часто тонкостенные паренхимные клетки сердцевины рано отмирают, заполняются воздухом, между ними возникают большие межклетники. В этом случае сердцевина становится белой и рыхлой, как, например, у бузины, кукурузы, подсолнечника.
Если сердцевина отмирает раньше, чем заканчивается рост междоузлий, то она разрывается и в центре стебля образуется полость. Такие полые стебли злаков называют соломинами. Полые (дудчатые) стебли характерны для зонтичных и тыквенных.
В отличие от корня в стебле первичного строения больший его объем занимает центральный цилиндр, а первичная кора относительно небольшая (особенно у однодольных). Как уже отмечалось при характеристике механических тканей, они занимают в корне и стебле неодинаковое положение в связи с разной нагрузкой, испытываемой этими органами.
Первичное строение стебля сохраняется в течение всей жизни побегов однодольных растений. У побегов хвойных голосеменных и двудольных покрытосеменных растений его можно наблюдать лишь на ранних этапах их развития. В связи с образованием и деятельностью вторичных латеральных меристем — камбия и феллогена — у этих растений происходит изменение первичного строения стебля на вторичное.
Образование годичных колец
Годичные кольца. Рассматривая древесину на поперечном срезе через многолетний стебель дерева, можно увидеть чередующиеся светлые и темные концентрические кольца. Каждая пара колец (светлое и темное) – это слой древесины, образовавшийся на протяжении одного года за счет деятельности камбия. Он называется годичным кольцом.
Весной клетки камбия активно делятся. Так возникают крупные клетки древесины, в частности, сосуды большого диаметра и с тонкими стенками. На поперечном срезе они составляют светлую часть годичного кольца. Летом же камбий формирует значительно более мелкие толстостенные сосуды и клетки механической ткани. Так возникает темный слой. Зимой клетки камбия вообще не делятся. Следующей весной его деятельность возобновляется и начинает формироваться новое годичное кольцо.
Годичные кольца могут «поведать» об условиях жизни растения. Так, их количество соответствует возрасту побега древесного растения. Ширина колец зависит от погодных условий в тот или иной год жизни растения. Больший прирост наблюдается в годы, благоприятные по своим погодным условиям. У старых деревьев активность камбия снижена, поэтому годичные кольца узкие, а прирост побегов незначительный.
Водоносная ткань
Клетки этой ткани отличаются большим запасом в вакуолях слизистых веществ, удерживающих влагу, что способствует удержанию и запасанию воды. Она хорошо развита у растений, приспособленных к жизни в засушливых местах с сухим климатом. Такие растения получили название — суккуленты: алоэ, кактусы. Как правило, они произрастают в местах с засушливым климатом.Водоносная паренхима при наступлении засухи постепенно отдает свои запасы воды другим, жизненно важным для растения тканям, в первую очередь хлорофиллоносной паренхиме.
ПП-проводящие пучки; Хл — хлорофилоносная ткань; Э — эпителий; ВТ — водоносная ткань
Вентиляционные ткани (аэренхима)
Аэренхима – это вентиляционная ткань или ткань проветривания. Главную функцию аэренхимы выполняют крупные межклетники, по которым и циркулирует воздух. Воздух необходим высшим растениям как для дыхания, так и для процессов фотосинтеза. Наличие аэренхимы характерно для водных или околоводных высших растений. Воздух, находящийся в системе полостей аэренхимы, не только вентилирует все части растения (в особенности подводные), но и придает им плавучесть, как, например, листьям кувшинки.
Аэренхима обычно имеет вид системы полостей с однослойными стенками. Клетки, слагающие стенки полостей могут иметь вытянутую форму или же могут быть шарообразной формы. Сами полости при этом в некоторых местах имеют тонкие пленчатые перегородки из одного ряда мелких клеток. Клетки этих перегородок имеют звездчатую форму, таким образом, между «лучей» данных клеток остаются мелкие отверстия в пленке (межклетники). Данные перегородки не мешают выполнять вентиляционную функцию аэренхиме, пропуская воздух через эти мелкие отверстия. Однако, если произойдет повреждение и полость начнет заполняться водой, то такая перегородка не попустит капельно-жидкую воду, поскольку поверхностное натяжение жидкости не позволит ей пройти сквозь мелкие отверстия. Такая аэренхима встречается у кувшинки, ириса, рдеста и т.д.
В другом случае аэренхима может быть целиком представлена только звездчатыми клетками. Такие клетки формируют трехмерную рыхлую ткань, похожую по консистенции на вату. Между «лучей» этих клеток также формируется одно большое общее межклеточное пространство, по которому циркулирует воздух. Такой тип аэренхимы характерен для ситников, осок, некоторых злаков и т.д. Также рыхлая аэренхима, многократно преломляя свет, придает белый цвет лепесткам некоторых растений.
Рисунок: Аэренхима. А – аэренхима на поперечном срезе стебля; Б – клетки пленчатой перегородки, разделяющей полости аэренхимы; В – аэренхима из трехмерно расположенных звездчатых клеток.
Вентиляционная ткань выполняет свою функцию за счет многочисленных увеличенных межклетников. Стоит помнить, что межклетники по типу происхождения делятся на три типа. Схизогенные межклетники образовались в результате простого расхождения клеток в пространстве. Лизигенные полости формируются в результате деградации (лизиса) некоторых клеток. Крупные рексигенные полости являются результатом механического разрыва тканей, например, в центре черешков или стеблей некоторых растений.
# Анатомия растений
# 10 класс
# 11 класс
Как образуется древесина?
Древесина – это одна из составляющих сосудисто-волокнистого пучка, она противопоставлена еще одной важной части пучка, формирующейся из того же прокамбия или камбия — лубу, или флоэме. В процессе формирования сосудисто-волокнистых пучков из прокамбия возможно два варианта событий:
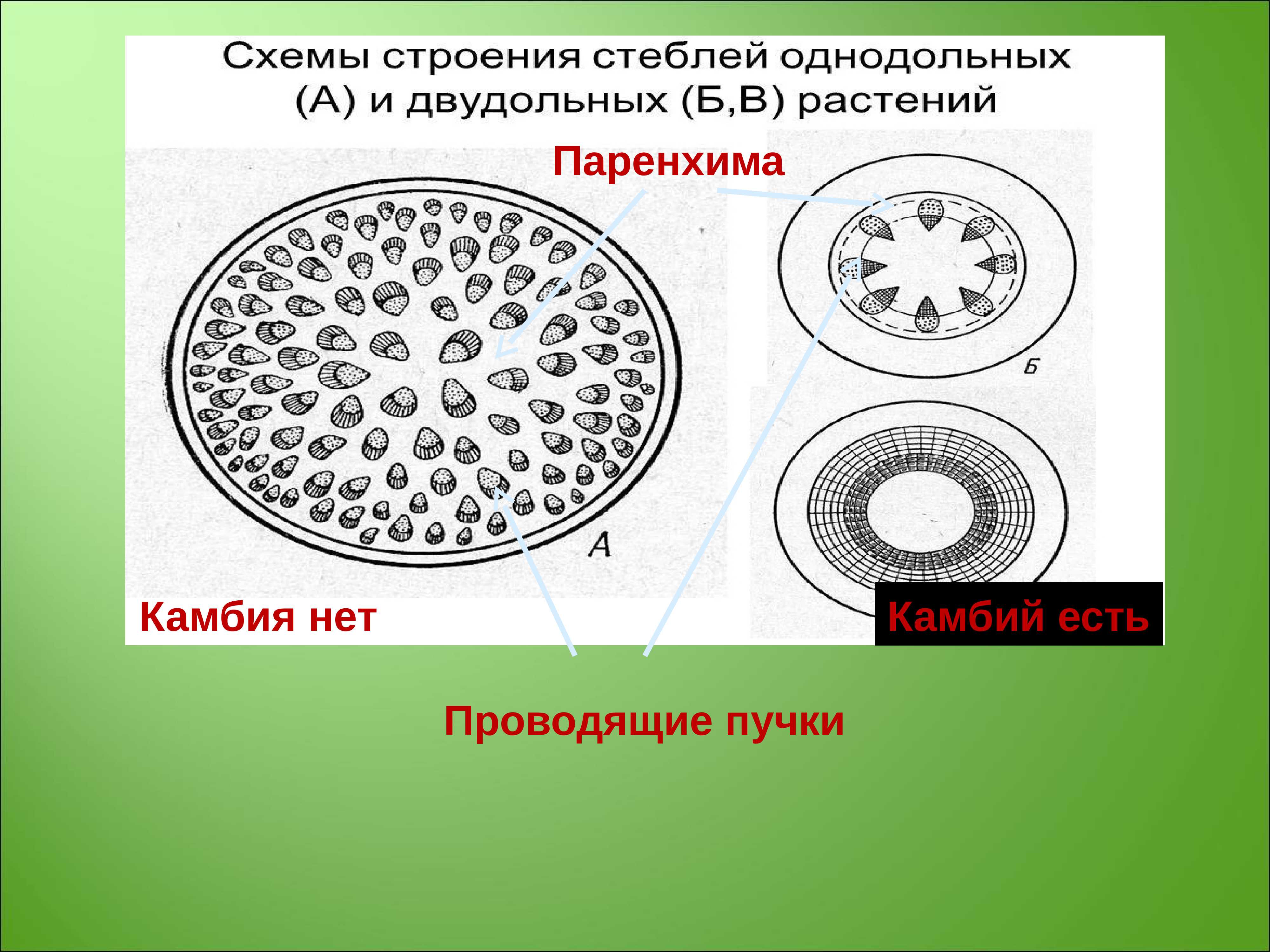
- все прокамбиальные клетки становятся элементами древесины и луба с образованием так называемых замкнутых пучков. Этот процесс характерен для высших споровых, однодольных и некоторых двудольных растений
- на границе между древесиной и лубом остаётся прослойка деятельной ткани, которую называют камбий. При этом формируются пучки открытые, что характерно для двудольных и голосеменных растений.
При первом варианте событий количество древесины не изменяется, и растение не может утолщаться. Если развитие идет по второму пути, то вследствие работы камбия ежегодно объем древесины увеличивается, и ствол растения медленно становится толще. У древесных пород российского региона древесина находится ближе к центру (оси) дерева, а луб — находится ближе к окружности (периферии). Ряд других растений имеет несколько иное взаимное расположение древесины и луба.



Именно деление клеток камбия в стебеле обеспечивает его рост в толщину. В процессе деления камбиальных клеток ? дочерних образующихся клеток отделяется в древесину, а ? – в луб. По этой причине прирост очень заметен в древесине. Камбий делится не равномерно, этот процесс зависит от сезона. В весенне – летний период деление активное, вследствие чего формируются крупные клетки, к осени деление замедляется, и формируются мелкие клетки. Зимой же камбий не делится. Таким образом, обеспечивается годичный прирост древесины, который хорошо заметен у многих деревьев, и называют его годичным кольцом. По количеству годичных колец специалисты вычисляют возраст побега и целого дерева.
Древесина имеет в своем составе уже отмершие клеточные элементы с одеревеневшими, главным образом, толстыми оболочками. Состав луба, напротив, представлен элементами живых клеток, с живой протоплазмой, клеточным соком и тонкой неодеревеневшей оболочкой. В тоже время, в лубе могут попадаться элементы мёртвые, толстостенные и одеревеневшие.
Обе составляющие сосудисто-волокнистого пучка имеют и еще одно физиологическое отличие. По древесине из земли к листьям движется сырой сок, который является водой с растворёнными в ней полезными веществами. А вот по лубу вниз течет пластический сок.
Процесс одеревенения клеточных оболочек характеризуется пропитыванием целлюлозной оболочки специальными веществами, которые объединены под общим названием лигнин. Наличие лигнина и вместе с тем одеревенение оболочки можно легко определить с помощью определенных реакций. Вследствие одеревенения, растительные оболочки растут в толщину, твердеют. В тоже время при лёгкой проницаемости для воды они утрачивают способности поглощать воду и разбухать.
Механическая ткань
1. КолленхимаКолленхима имеет неравномерно утолщенные клеточные стенки, в основе которых находятся полисахариды: целлюлоза, гемицеллюлозы. Способна к фотосинтезу, тк клетки содержат хлорофилл (в подземных частях растения колленхима не встречается).Классификация: 1. Уголковая колленхимаКлетки в виде шестиугольников, клеточная стенка их утолщена в углах, а между углами стенки тоньше, поэтому данная ткань относится к неравномерно утолщенным. Встречается в стеблях щавеля, гречихи, тыквы — двудольных растений, в крупных жилках листа, черешках листьев.2. Пластинчатая колленхимаХарактерна для молодых стеблей многих деревьев. В отличие от уголковой колленхимы клетки имеют форму параллелепипеда, вытянуты параллельно поверхности стебля, их наружные и внутренние стенки утолщены.3. Рыхлая На раннем этапе развития клетки данной ткани разъединяются в углах с последующим образованием межклетников (пространства в тканях растения), имеются в стеблях красавки, мать-и- мачехи, горца земноводного.
1 — уголковая; 2 — пластинчатая; 3 — рыхлая колленхима
2. СклеренхимаСостоит в основном из мертвых клеток, встречается в органах высших растений, выдерживает большие нагрузки. Клеточная стенка пропитана лигнином — смесь ароматических полимеров. Ядро и цитоплазма клеток разрушаются. Склеренхима представлена двумя типами тканей: Представлены вытянутыми и заостренными клетками, форма которых называется «прозенхимная». Клетки плотно прилежат друг к другу, их оболочка очень прочная, клеточные стенки утолщены равномерно. Волокна встречаются во всех органах растения в виде тяжей, могут быть рассеянны в проводящей ткани, собираться в группы или идти сплошным цилиндрическим кольцом.
В текстильной промышленности широко используются не одревесневшие лубяные волокна, к примеру — льна. Из них получают разные ткани, широко применяемые в быту. Так что обязательно отметьте их хозяйственное значение.
Стенки этих клеток сильно одревесневшие, могут быть пропитаны кремнеземом, известью, кутином. В случае, если диаметр клеток одинаковый (плоды груши) их также называют каменистые клетки (брахисклереиды). Палочковидные склереиды встречаются в семенах бобовых. Остеосклереиды имеют расширение на обоих концах клетки, встречаются в листьях чая. В листьях камелии cклереиды приобретают удивительную форму, напоминающую звезду, они называются астросклереидами.Как вы уже убедились, склереиды представляют собой мертвые клетки самых различных форм, обнаруживаются во многих органах растения.
Анатомия стебля
Основные функции стебля — проведение воды и питательных веществ. Дополнительными функциями могут быть фотосинтез, обычно происходящий в молодых стеблях растений, и отложение запасных питательных веществ в многолетних стеблях.
Анатомическое строение стебля растений обусловлено его главными функциями. Для него характерно развитие механической и проводящей тканей растений. Кроме того, для стебля характерна сложная система меристем — верхушечные, боковые и вставочные, определяющие нарастание его в течение длительного времени и возникновение новых органов.
Стебель растения возникает из апикальной меристемы, из которой дифференцируются три слоя тканей: покровная, проводящая, основная. Сначала эти ткани представлены так называемой первичной меристемой (промеристемой), состоящей из протодермы, прокамбия и основной меристемы.
Первичные проводящие ткани развиваются из прокамбия. Первые элементы флоэмы дифференцируются из наружных, расположенных к периферии клеток прокамбия. Первичная флоэма представлена тонкостенными недолговечными удлиненными клетками и носит название протофлоэмы, а наружные клетки ее могут быть представлены механическими волокнами.
Первичные элементы ксилемы — трахеиды, реже сосуды с кольчатыми и спиральными утолщениями стенок — возникают позже из внутренних клеток прокамбия и определяются в целом как протоксилема. В ее состав помимо проводящих элементов входят паренхимные клетки.
Позже внутрь от протофлоэмы дифференцируется имеющая более или менее типичное для флоэмы строение. Наружу от протоксилемы формируется метаксилема, состоящая из трахеид или трахей с более утолщенными одревесневшими стенками.
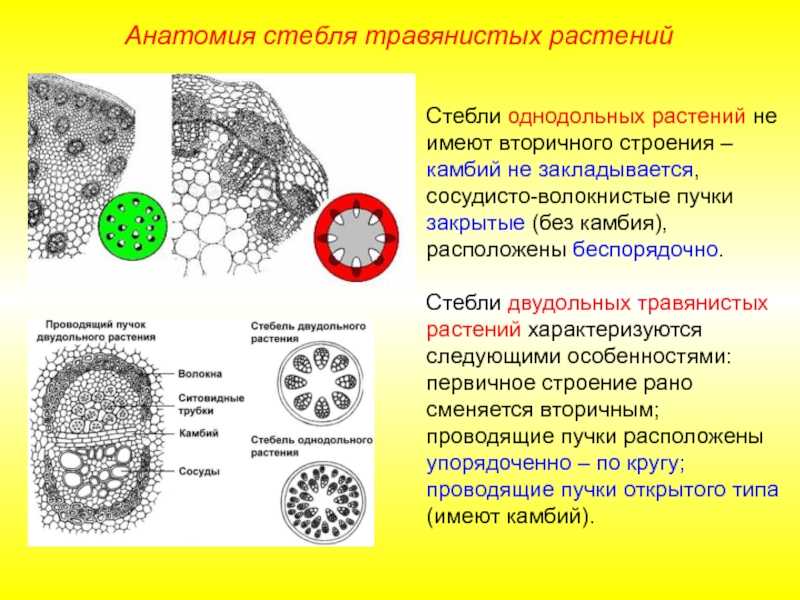
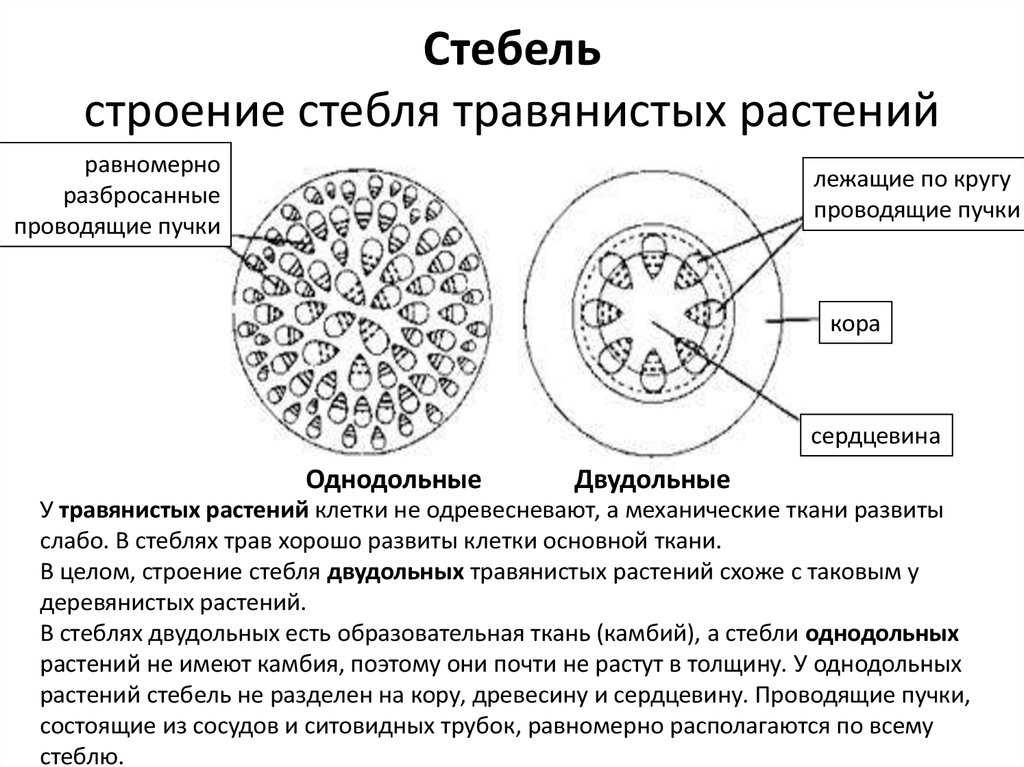
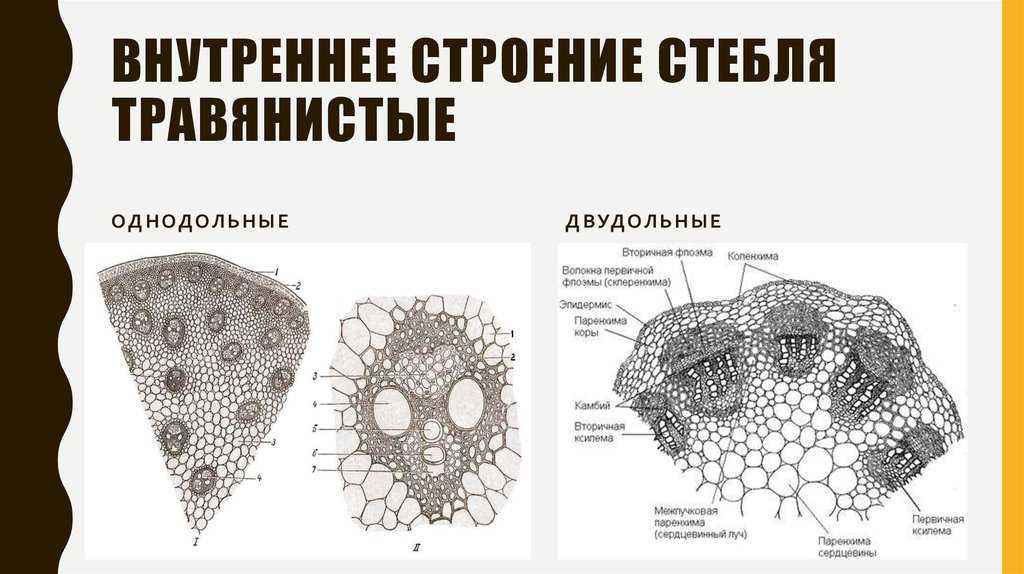
Таким образом, за счет деятельности прокамбия и остальной меристемы апекса возникает первичное строение стебля растения. У однодольных растений весь прокамбий дифференцируется в элементы первичных проводящих тканей. Стебли однодольных, особенно травянистых (злаков), по сравнению со стеблями двудольных растений имеют более простое строение, для них характерно в основном первичное строение.
У двудольных растений в средней части прокамбиального тяжа происходит образование камбия и начинается образование вторичных проводящих тканей (метафлоэмы и метаксилемы), объем которых увеличивается за счет деления клеток камбия.
Пучки, состоящие только из первичных тканей, как у однодольных, закрытые-, пучки с камбием, характерные для двудольных, — открытые.
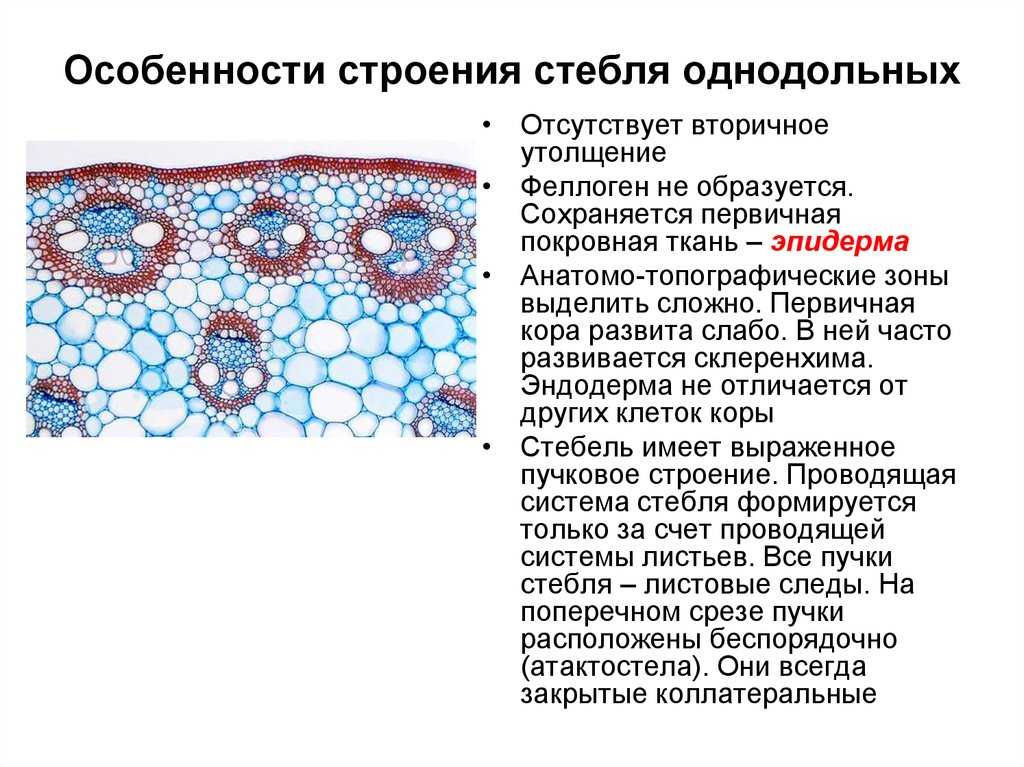
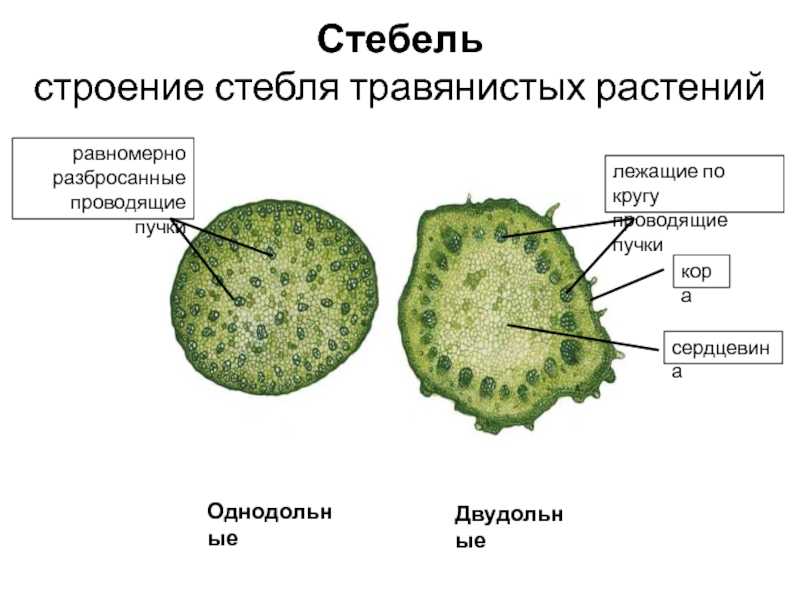
Для травянистых однодольных растений характерно диффузное распределение проводящих пучков. Проводящие пучки закрытые, коллатеральные, реже концентрические. Из механических тканей наиболее развита склеренхима, колленхима встречается у немногих растений. Вторичного утолщения у травянистых однодольных нет.
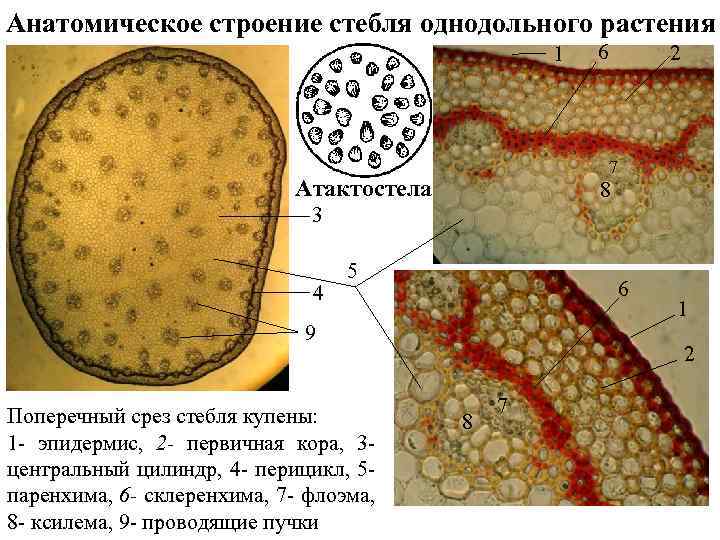
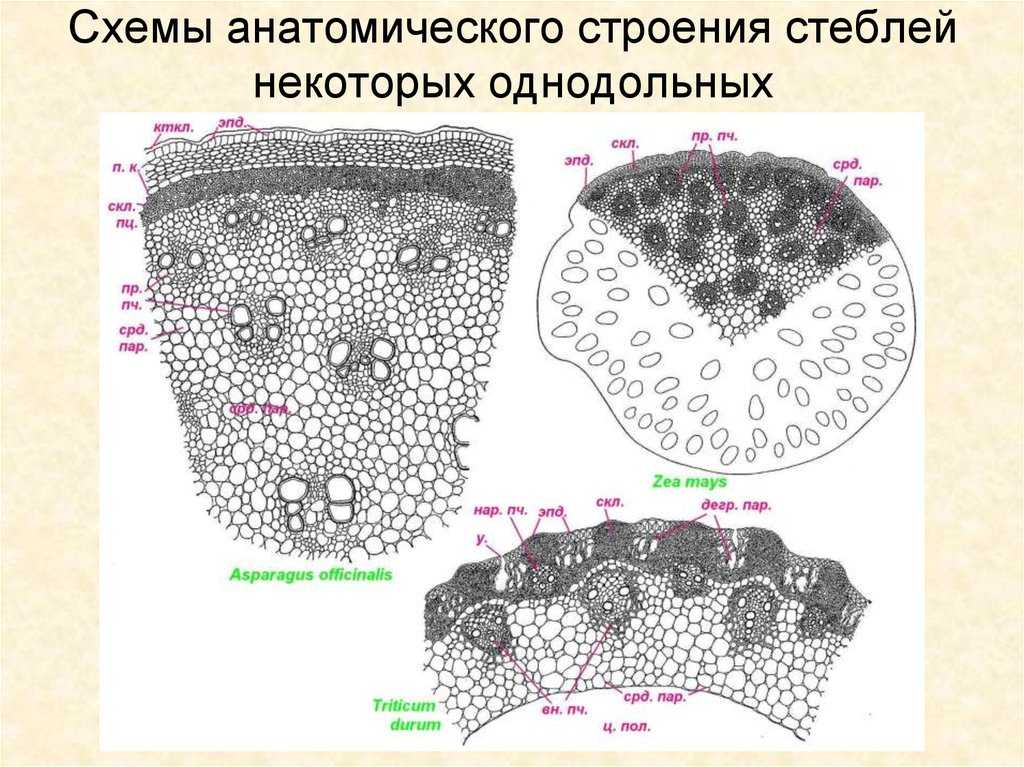
Первичное строение стебля однодольных можно рассмотреть на примере кукурузы. С поверхности стебель покрыт однослойной эпидермой со слоем кутикулы. Под эпидермой расположен слой паренхимы в виде сплошного кольца. Клетки паренхимы непосредственно под эпидермой склерифицированы, к центру крупные, тонкостенные. В молодых стеблях под эпидермой размещается хлорофиллоносная паренхима. С возрастом утолщается склерифицированное кольцо, стенки клеток древеснеют и образуется резкая граница между этими клетками и клетками обычной паренхимы.
Клетки паренхимы крупные, с тонкими стенками, между ними — межклетники. К концу вегетации большая часть клеток паренхимы заполняется воздухом, сердцевина становится белой, мягкой. В паренхимной ткани беспорядочно разбросаны закрытые сосудисто-волокнистые пучки коллатерального типа.
На поперечных срезах пучков флоэма имеет вид сеточки, крупные ячеи ее соответствуют поперечному сечению ситовидных трубок. В узлах сеточки расположены мелкие квадратные или прямоугольные сопровождающие клетки. Во взрослых стеблях функционирует только метафлоэма. Протофлоэма, занимающая в пучке периферическое положение, обычно сжата и деформирована. Ксилема представлена 3-5 сосудами. Внутрь от метафлоэмы находятся два очень крупных пористых сосуда метаксилемы, соединенных механическими элементами. Между сосудами метаксилемы расположены узкопросветные сосуды протоксилемы с кольчатыми и спиральными утолщениями оболочек. При развитии пучка некоторые элементы протоксилемы разрушаются и на их месте возникает воздушная полость, ограниченная паренхимными клетками.
Механические ткани
Существует две специализированные механические ткани высших растений – склеренхима и колленхима.
Склеренхима, как правило, состоит из клеток вытянутой формы – волокнообразных. Их клеточные стенки утолщаются и лигнифицируются, то есть одревесневают. Живое содержимое клетки впоследствии отмирает. Таким образом, склеренхима – это мертвая ткань, механическую функцию в которой выполняют жесткие клеточные стенки. Склеренхима твердая жесткая ткань и в растении она выполняет армирующую функцию, располагаясь обычно тяжами или слоями. Однако иногда склеренхима может быть представлена в виде отдельных клеток с одревесневшими клеточными стенками, разбросанных в толще некой мягкой ткани (например, паренхимы). Такие клетки называются склереидами. По форме различают разные типы склереид: брахисклереиды, астросклереиды, остеосклереиды и волокнистые склереиды. Все склеренхимные элементы вместе составляют стереом – совокупность всех толстостенных одревесневших клеток растения. Следует также помнить, что отчасти механическую функцию, подобно склеренхиме, выполняет водопроводящая ткань ксилема (в особенности ядровая древесина – вторичная ксилема, прекратившая проводить воду).
Рисунок 1: Склеренхима.
Колленхима также является механической тканью, однако клетки ее остаются живыми. Их клеточные стенки утолщаются, но неравномерно и не одревесневают. Живые клетки упругие, так как находятся под тургорным давлением, а клеточные стенки эластичны, поскольку состоят из полисахаридов. Именно эти свойства и позволяют колленхиме выполнять свою механическую функцию. Таким образом, колленхима – это живая упругая эластичная механическая ткань. Обычно колленхима располагается в тех органах высших растений, которые подвержены изгибу и должны быть упругими. Например, это стебли травянистых растений, особенно если стебель граненый или ребристый, то вдоль граней под эпидермой, скорее всего, располагаются тяжи колленхимы. Также колленхима часто встречается в листьях в черешке и вдоль средней жилки, поскольку именно эти части должны быть эластичными и упругими. Выделяют три типа колленхимы: уголковую (клеточные стенки утолщены в местах контакта трех и более клеток – «в уголках»), пластинчатую (утолщены продольные клеточные стенки) и рыхлую (похожа на уголковую, но с крупными межклетниками).
Рисунок 2: Колленхима. А – рыхлая; Б – пластинчатая; В – уголковая. 1 – первичная; клеточная стенка; 2 – вторичная клеточная стенка; 3 – межклетник; 4 – протопласт.


Проводящие ткани (ксилема, флоэма)
Ксилема – сложная ткань, то есть состоит из клеток разной морфологии. В состав ксилемы одновременно входят и проводящие, и механические, и запасающие элементы.
Ксилема проводит воду с растворенными в ней минеральными веществами от корней по всему остальному телу растения. Таким образом, по ксилеме в основном осуществляется восходящий ток. Проводящие элементы ксилемы – это сосуды и трахеиды. Следует помнить, что ксилема голосеменных растений лишена сосудов. Трахеида образуется из клетки удлиненной формы, ее клеточная стенка утолщается и лигнифицируется, то есть одревесневает. Протопласт при этом отмирает и в результате получается мелкий капилляр, по которому может транспортироваться вода. Прочные клеточные стенки предохраняют просвет капилляра от схлопывания. От трахеиды к трахеиде вода транспортируется через специальные поры. Сосуд, по сути, является таким же капилляром, как и трахеида, но более длинным, широкопросветным и многоклеточным. Каждый сосуд состоит из отдельных клеток (члеников сосуда) с одревесневшей оболочкой и отмершим протопластом, между члениками сосуда формируются уже не поры, а перфорационные пластинки (то есть сквозные отверстия). Между сосудами, как и между трахеидами, есть поры, через которые также может транспортироваться вода. Кроме проводящих элементов, в состав ксилемы входят механические волокна – волокна либриформа. Это удлиненные клетки, похожие на трахеиды, однако их клеточные стенки очень сильно утолщены и лигнифицированы. Просвет таких капилляров слишком мал для осуществления транспорта воды, зато толстая и прочная клеточная стенка выполняет механическую функцию подобно склеренхиме. Ксилема в основном состоит из мертвых клеток, обычно небольшой процент живых клеток представлен древесинной паренхимой. Эти клетки в основном выполняют запасающую функцию.
Флоэма, как и ксилема, – это сложная ткань, которая состоит из разных клеток. В состав флоэмы входят проводящие механические и паренхимные (в том числе запасающие) элементы.
Флоэма транспортирует раствор питательных веществ, в основном это углеводы, образовавшиеся в результате фотосинтеза. Поскольку фотосинтез происходит преимущественно в листьях, а питательные вещества нужно доставлять во все части растения, в том числе и в корни, по флоэме преимущественно осуществляется нисходящий ток веществ. Проводящими элементами являются ситовидные клетки. Это живые клетки, они имеют вытянутую форму, а в их стенках формируются так называемые ситовидные поля. Ситовидное поле – это участок клеточной стенки, где близко друг к другу расположено множество плазмодесм. Через ситовидные поля происходит транспорт веществ от одной ситовидной клетки к другой. У покрытосеменных растений проводящими элементами флоэмы являются ситовидные трубки. Ситовидная трубка – это более длинная многоклеточная проводящая структура. Состоит она из одного ряда клеток, называемых члениками ситовидной трубки. В местах контакта члеников друг с другом формируются ситовидные пластинки – участки клеточной стенки, где расположено одно или несколько сближенных ситовидных полей. Вещества транспортируются по внутреннему содержимому живой клетки. Однако в ситовидных элементах деградируют многие органеллы, в том числе и ядро. Таким образом, ситовидная клетка и членик ситовидной трубки находятся в «полуживом» состоянии. При этом существуют специальные клетки, которые поддерживают ситовидные элементы в этом состоянии, обеспечивают и регулируют их жизнедеятельность. Такие клетки называются клетками-спутницами у члеников ситовидных трубок, а ситовидные клетки поддерживают специальные клетки Страсбургера. Кроме проводящих элементов во флоэме, как и в ксилеме, находятся паренхимные (запасающие) клетки, а также механические элементы (лубяные волокна). Волокна обычно представлены удлиненными клетками с толстой одревесневшей клеточной стенкой.
Рисунок: Проводящие ткани. А – ксилема; Б – флоэма. 1 – сосуды ксилемы; 2 – трахеиды; 3 – клетки древесной паренхимы; 4 – поры; 5 — ситовидные трубки; 6 – клетки – спутницы; 7 – ситовидные поля; 8 – клетки лубяной паренхимы.
Поглощающие ткани
Высшие растения поглощают воду с помощью специальных тканей. У мохообразных отсутствуют корни, и всасывание воды происходит всей поверхностью тела (например, с помощью гиалиновых клеток у сфагновых мхов) или с помощью ризоидов – длинных тонкостенных клеток. Сосудистые растения имеют корни, поверхность которых покрыта ризодермой (эпиблемой) – специализированной всасывающей тканью. Ризодерма гомологична эпидерме, то есть также формируется из одного внешнего слоя клеток, покрывающих орган. Однако ризодерма не является покровной тканью, поскольку практически не выполняет защитную функцию. Ее клетки тонкостенные и специализируются на поглощении воды и минеральных солей из почвы, поглощение при этом происходит избирательно и с затратой энергии. В ризодерме различают два типа клеток: трихобласты и атрихобласты. У трихобластов наружная часть клетки выпячивается и образует длинный вырост – корневой волосок, служащий для увеличения поверхности всасывания. Корневой волосок выделяет слизь, которая помогает растворять поглощать минеральные вещества из почвы. Атрихобласты не формируют корневых волосков, но также поглощают вещества своей поверхностью.
Рисунок: Ризодерма. А – Продольный разрез корня; Б – Клетки ризодермы. 1 – зона проведения; 2 – зона всасывания; 3 – зона роста; 4 – зона деления; 5 – корневые волоски; 6 – корневой чехлик.
У некоторых тропических эпифитных растений вместо ризодермы развивается веламен. Веламен гомологичен ризодерме, но в отличие от нее является многослойной тканью и состоит из отмерших клеток. Их клеточные стенки имеют спиральные утолщения, которые служат ребрами жесткости, сами клеточные стенки частично разрушаются, а внутреннее содержимое клеток отмирает. В результате получается структура наподобие губки, которая способна впитывать воду из влажного воздуха, тумана или осадков. Таким образом, веламен поглощает вещества пассивно и не избирательно. Направленный и избирательный транспорт воды дальше внутрь корня происходит при участии экзодермы, подстилающей веламен (как, впрочем, и любую ризодерму).
Основная ткань
Здесь идет газообмен с окружающей средой, фотосинтез, запасание питательных веществ, запасание воды. Составляет основную часть массы растения, состоит из живых паренхиматозных клеток, образованных из первичной меристемы — верхушечной (апикальной).
Строение листа
Ассимиляционная ткань (хлоренхима)Ассимиляционная = синтезирующая. За счет содержания хлоропластов и хлорофилла в данной ткани, здесь активно идет процесс фотосинтеза. Например, это столбчатая ткань мякоти листа, или мезофилл — мягкая ткань, заключенная между двумя слоями эпидермиса в листьях растений.Хлоренхима расположена непосредственно под эпидермисом, это обеспечивает ее хорошее освещение и газообмен с окружающей средой. Она встречается в надземных органах растений, таких как листья, молодые побеги. Но это не исключает возможность ее возникновения на освещенных корнях, к примеру, в корнях водных растений, воздушных корнях.
Воздухоносная ткань (аэренхима)Главная ее функция — газообмен. Отличается наличием межклетников — тканевых пространств, служащих вместилищем для газов. Сквозь устьица воздух межклетников путем диффузии уравнивается по составу с атмосферным воздухом. В межклетниках из атмосферного воздуха клетки растения поглощают углекислый газ и выделяют в полость кислород, который затем поступает в окружающую среду. Локализована в губчатой ткани листа.Благодаря наличию межклетников в ткани ее удельный вес уменьшается, и она помогает листьям водоплавающих растений держаться на плаву.