Содержание
-
Слайд 1
Анализирующее скрещивание Взаимодействие аллельных генов Множественный аллелизм
-
Слайд 2
Анализирующее скрещивание
Анализирующее скрещивание – это скрещивание особи с неопределенным генотипом (АА или Аа) с рецессивной гомозиготной особью (аа).
Если в потомстве от такого скрещивания не наблюдается расщепления, т.е. все особи фенотипически одинаковы (Аа), то исследуемая особь гомозиготна (АА);Если образуется неоднородное потомство в соотношении 50/50 (1Аа:1 аа), то исследуемая особь была гетерозиготной (Аа).
1 случай:
Р: ААхаа;
G: А а;
F1: все Аа;1 случай:
Р: Аахаа;
G: А и а а;
F1: 50% Аа и 50% аа. -
Слайд 3
1.Доминирование
2.Неполное доминирование
3. Сверхдоминирование
4. Кодоминирование
-
Слайд 4
Взаимодействие аллельных генов
1.Доминирование.Одна аллель, называемая доминантной, полностью скрывает присутствие другой аллели (рецессивной), поэтому гетерозиготаАа имеет признаки доминантного аллеля (опыты Менделя с горохом).
-
Слайд 5
-
Слайд 6
Взаимодействие аллельных генов
2.Неполное доминирование
У гибрида F1 признак носит промежуточный характер, т.к. доминантная аллель не полностью скрывает рецессивную, и она частично проявляется (опыты Менделя с ночной красавицей).
А – пурпурная окраска;
а – белая окраска;
Аа – розовая окраска.Расщепление у гибридов F2для неполного доминирования1:2:1:
-
Слайд 7
-
Слайд 8
Взаимодействие аллельных генов
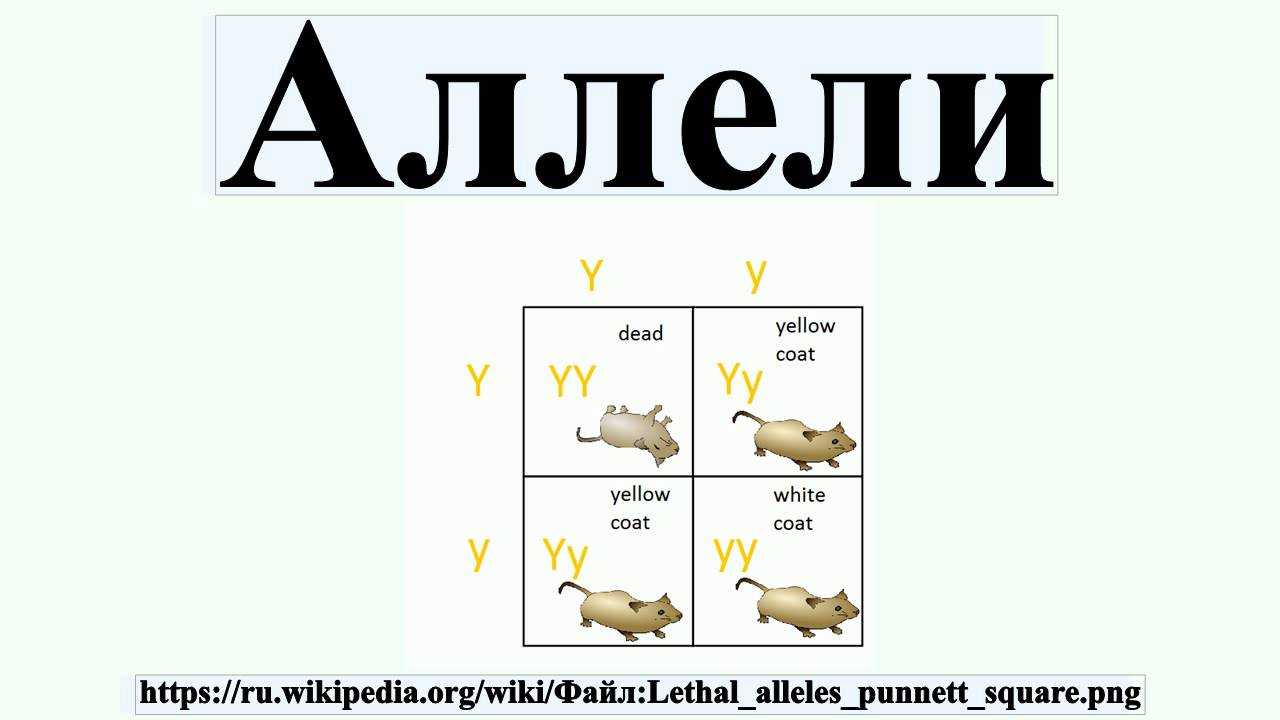
3. Сверхдоминирование – явление, при котором гетерозигота обладает бóльшей жизнеспособностью по сравнению с обеими гомозиготами по данной паре аллелей.
Например, у дрозофилы известна рецессивная летальная мутация, гетерозиготы по которой обладают бóльшей жизнеспособностью (плодовитостью, продолжительностью жизни), чем доминантная гомозигота дикого типа.
Серповидно-клеточную анемию можно также рассматривать как явление сверхдоминирования, так как у гетерозигот есть преимущество – устойчивость к малярии.
Явление гибридной силы – гетерозиса многие ученые объясняют взаимодействием генов по типу сверхдоминирования.
-
Слайд 9
Множественный аллелизм
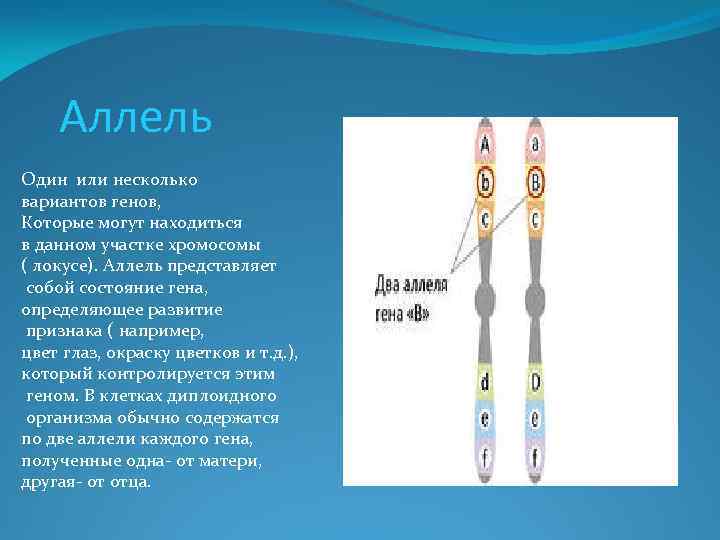
У одного организма может присутствовать только 2 аллеля одного гена — Аа (по одному в каждой гомологичной хромосоме). Однако, ген мутирует неоднократно, и в популяции новые аллели накапливаются. Из таких аллелей можно составить серию со сложными отношениями между собой.
А) Например, у кроликов известна серия аллелей по окраске шерсти. Ген А (черная окраска) доминирует над всеми, ген аш(серая) по отношению к гена аг (гималайская) и а (белая) ведет себя как доминантный, а ген агдоминирует над геном а.
Таким образом, А > аш> аг> а.
Интересно отметить, что кролики с генотипами ашаги аша имеют промежуточную между шиншилловым и белым – светло-серую — окраску. Здесь проявляется неполное доминирование аллели аш над аллелями аги а.
-
Слайд 10
-
Слайд 11
Б). У мухи дрозофилы известна серия аллелей по цвету глаз, состоящая из 12 типов: красная, коралловая, вишневая, абрикосовая и т.д. до белой, определяемой рецессивным геном.
В). Другим примером множественного аллелизма может служить система групп крови АВО, открытая Карлом Ландштейнером в 1900 году. Существуют 4 группы крови в системе АВО: О, А, В, АВ. Они определяются тремя аллелями одного гена: IА, IВ, i. Аллели IА и IВ доминантны по отношению к аллели i, но кодоминантны по отношению друг к другу.
-
Слайд 12
Взаимодействие аллельных генов
4. Кодоминирование-проявление в гетерозиготном состоянии признаков, определяемых обеими аллелями: например, каждый из отдельных генов кодирует определенный белок, и у гетерозиготного организма синтезируются оба. Например, когда один из родителей имеет группу крови А, а второй группу крови В, то в крови их детей присутствуют белки (антигены), характерные для обеих этих групп, что приводит к образованию группы крови АВ.
IA
IA
IB
IB
i
i
Посмотреть все слайды
Что такое файл cookie и другие похожие технологии
Файл cookie представляет собой небольшой текстовый файл, сохраняемый на вашем компьютере, смартфоне или другом устройстве, которое Вы используете для посещения интернет-сайтов.
Некоторые посещаемые Вами страницы могут также собирать информацию, используя пиксельные тэги и веб-маяки, представляющие собой электронные изображения, называемые одно-пиксельными (1×1) или пустыми GIF-изображениями.
Файлы cookie могут размещаться на вашем устройстве нами («собственные» файлы cookie) или другими операторами (файлы cookie «третьих лиц»).
Мы используем два вида файлов cookie на сайте: «cookie сессии» и «постоянные cookie». Cookie сессии — это временные файлы, которые остаются на устройстве пока вы не покинете сайт. Постоянные cookie остаются на устройстве в течение длительного времени или пока вы вручную не удалите их (как долго cookie останется на вашем устройстве будет зависеть от продолжительности или «времени жизни» конкретного файла и настройки вашего браузера).
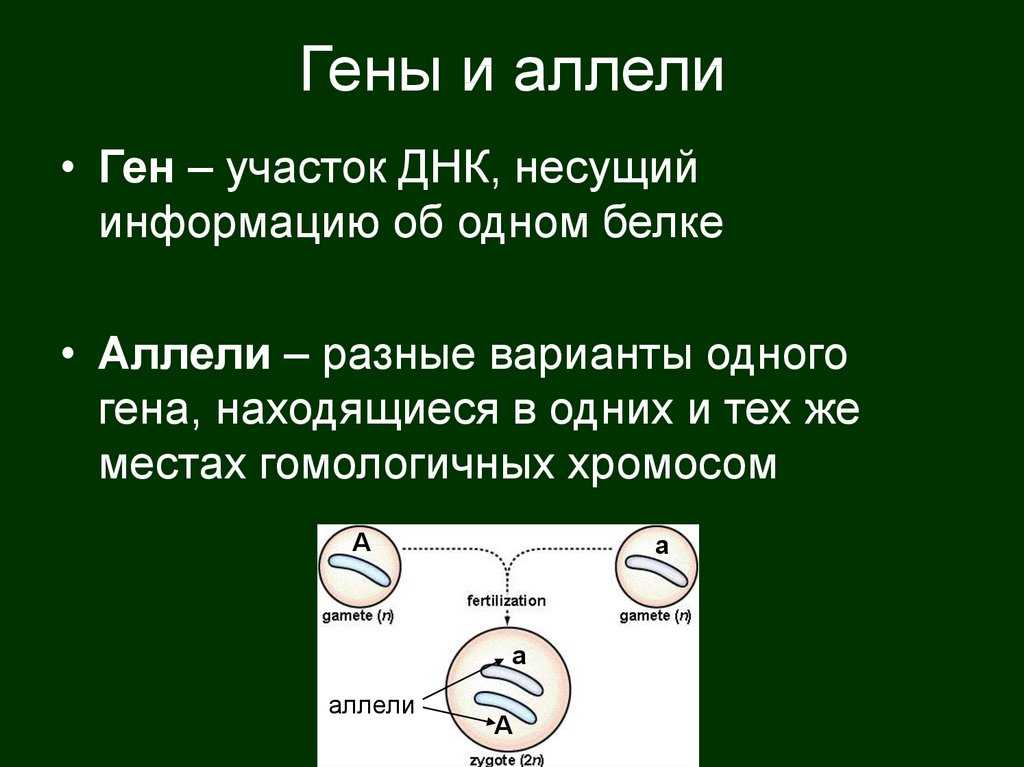
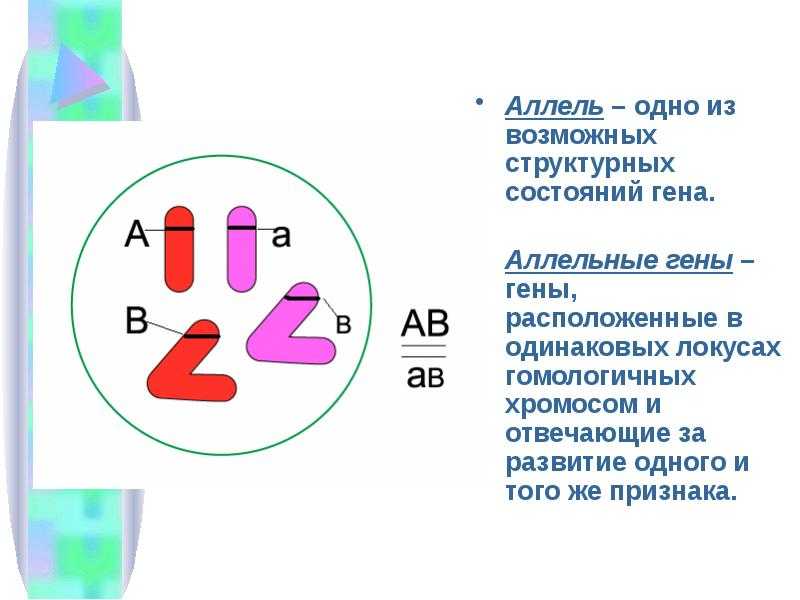
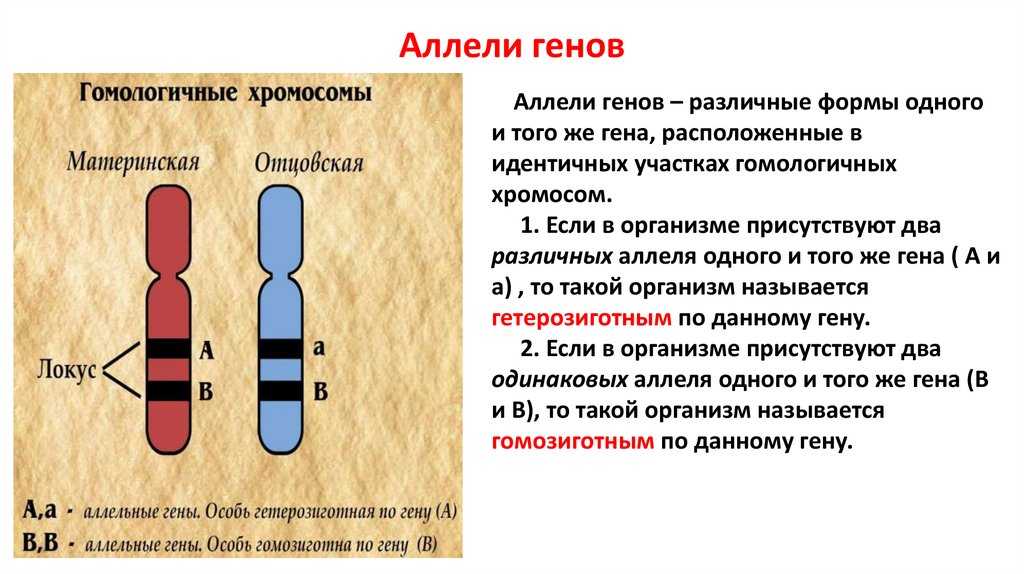
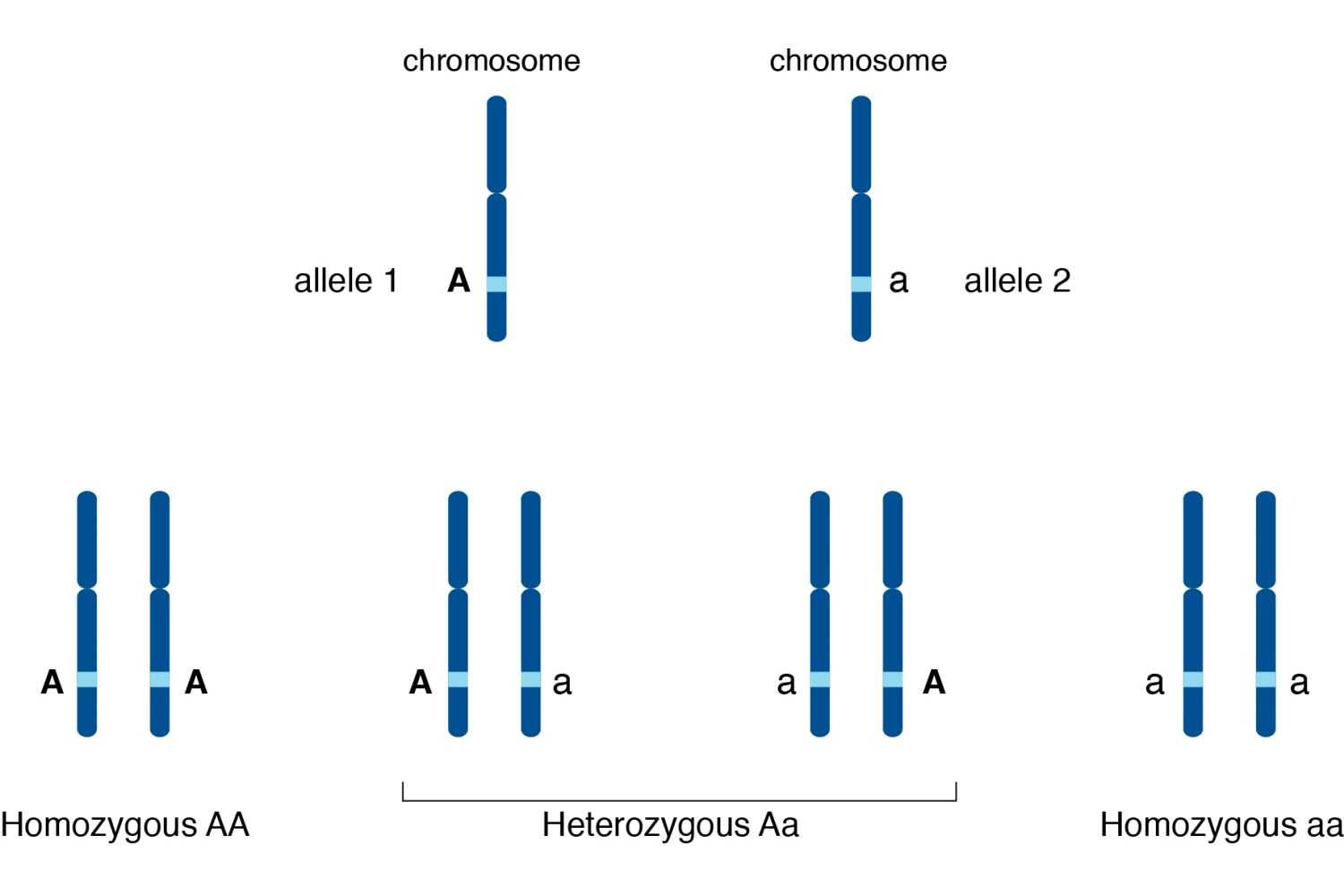
Взаимодействие аллельных генов
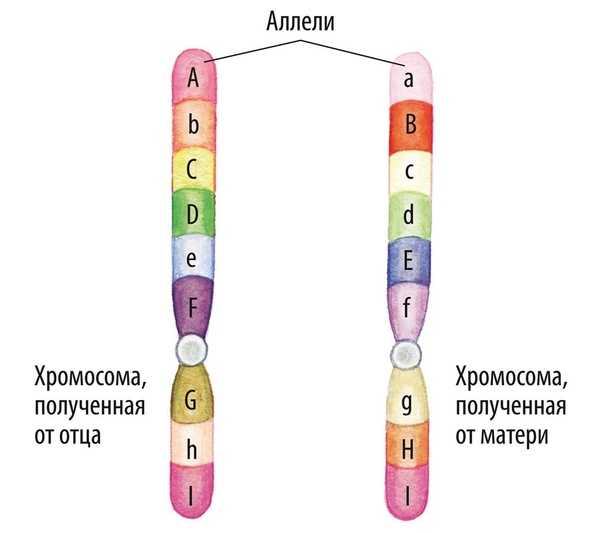
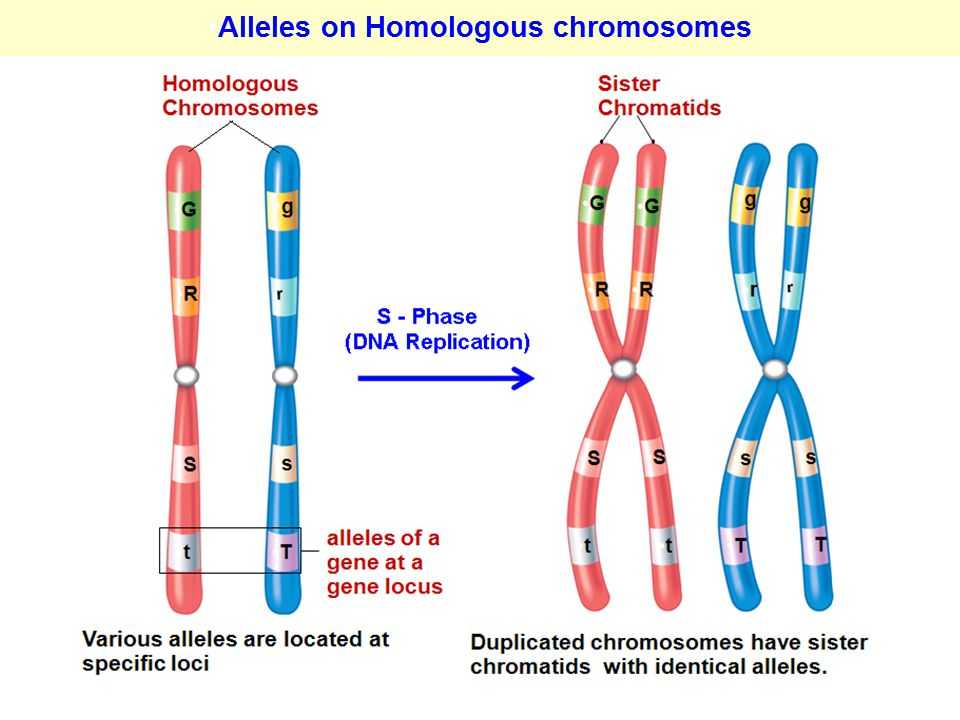
Аллели одного гена, находясь в одном организме, взаимодействуют между собой, и от этого взаимодействия зависит, как проявит себя признак, обусловленный соответствующим геном.
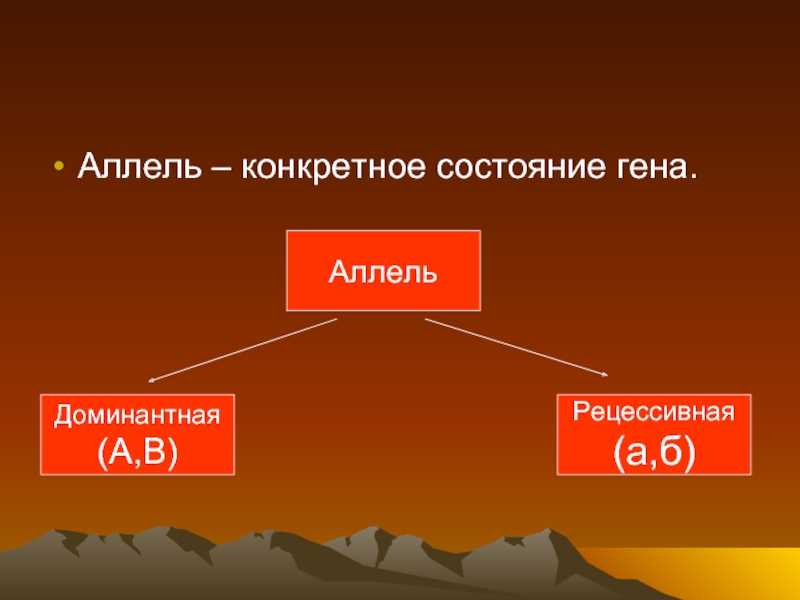
Полное доминирование
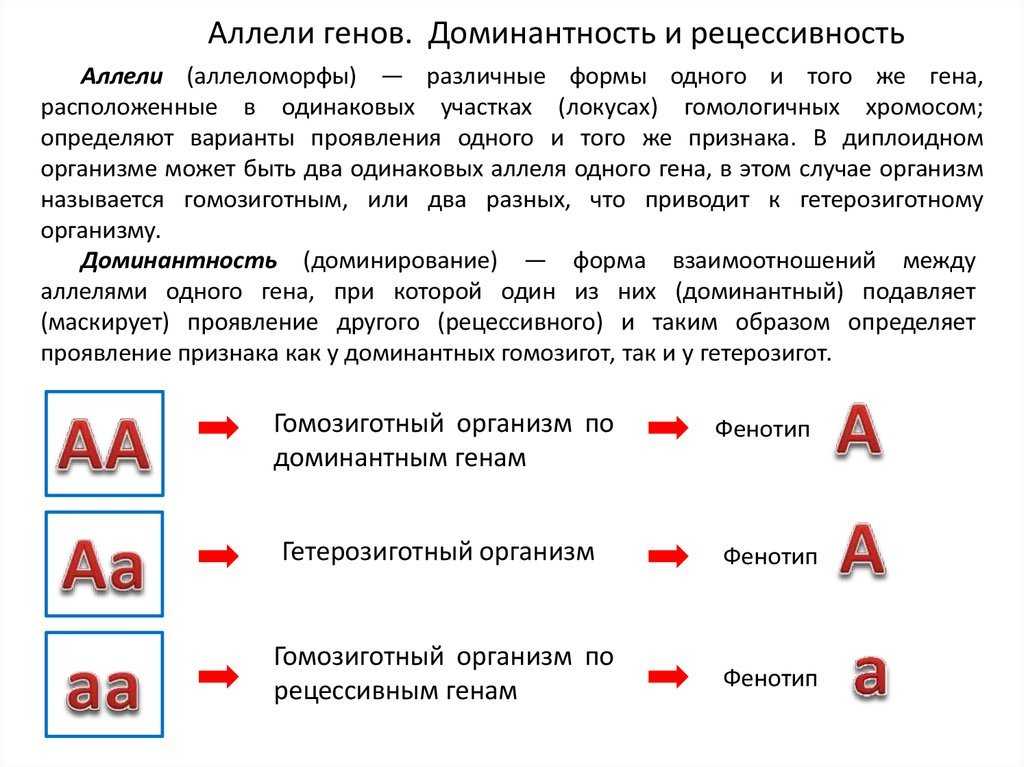
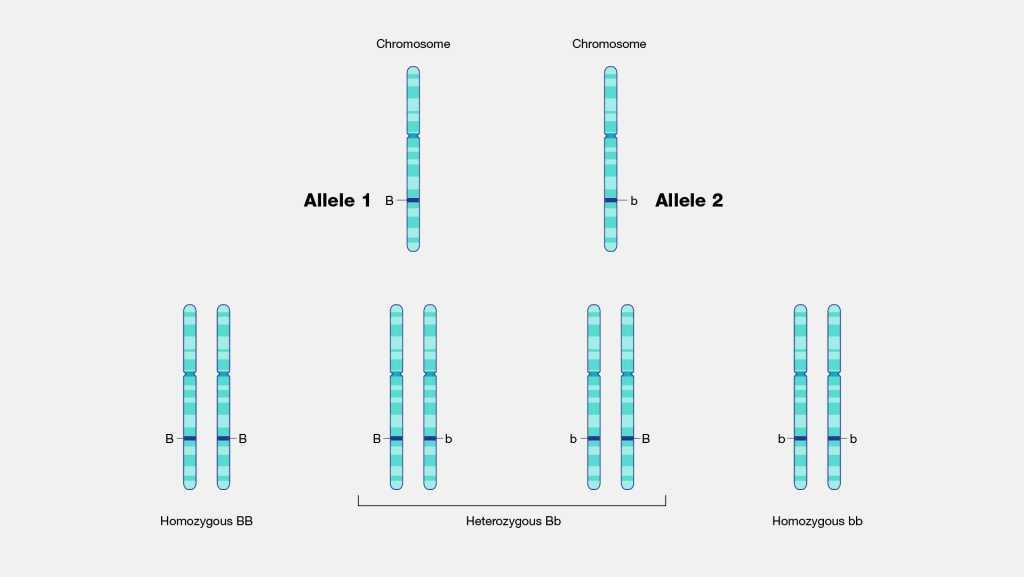
Чаще всего встречается такой тип взаимодействия как полное доминирование, при котором один аллельный ген проявляется и полностью подавляет проявление другого аллельного гена. В данном случае первый называется доминантным, а второй — рецессивным.
В генетике принято доминантный ген обозначать большой латинской буквой (например, ), а рецессивный — малой (). Если особь гетерозиготна, то ее генотип будет . Если гомозиготна, то или .
В случае полного доминирования генотипы и будут иметь одинаковый фенотип.
Полное доминирование наблюдается при проявлении, например, цвета семян гороха. У гетерозиготных () растений по этому признаку семена всегда желтые, как и у гомозиготных по доминантному аллелю (). Зеленый цвет определяется рецессивным аллелем и поэтому подавляется.
При множественном аллелизме отношения между аллелями могут быть неоднозначными. Если имеет место быть исключительно полное доминирование, то один ген может быть доминантным по отношению ко второму, но рецессивным по отношению к третьему. В таком случае строят ряды ( > > > …), в которых отражают отношения доминирования. Так, например, наследуется окраска шерсти у многих животных, цвет глаз.
Кроме полного доминирования встречаются и другие типы взаимодействия аллелей: неполное доминирование, кодоминирование, сверхдоминирование, комплементация аллелей и некоторые другие.
Неполное доминирование
В случае неполного доминирования гетерозиготный генотип будет иметь промежуточное значение признака. В этом случае доминантный аллель лишь частично подавляет рецессивный. В результате в фенотипе проявляется некое промежуточное значение признака.
Например, цвет цветков ночной красавицы с генотипом красный, — белый, — розовый.
При неполном доминировании значение признака не обязательно будет ровно «по середине». Чаще бывает отклонение в сторону доминантного, то есть когда фенотип генотипа хоть и не сходен с , но ближе к нему, чем к фенотипу генотипа .



Кодоминирование
При кодоминировании два аллельных гена, оказавшись в одном организме, полностью проявляют себя. Ни один из них никак не подавляет другой. В случае кодоминирования отсутствуют доминантно-рецессивные отношения между аллелями. В итоге в организме синтезируется два разных белка, определяющих один и тот же признак. Нечто похожее происходит при сверхдоминировании и межаллельной комплиментации.
При множественном аллелизме в одной паре аллелей может быть отношение полного доминирования, а в другой — кодоминирования. Так, например, обстоит дело с группами крови человека. Аллель является рецессивным и подавляется доминантными аллелями и . Если организм , то у него первая группа крови. Если или — вторая, или — третья. Если же человек имеет аллели и , то есть генотип , то у него четвертая группа крови, при которой в крови присутствуют два антигена (за наличие одного отвечает аллель , за наличие другого — ).
Сверхдоминирование
Сверхдоминирование — это такой тип взаимодействия аллельных генов, когда у гетерозиготных особей () признак сильнее проявляется, чем у обеих гомозиготных ( и ). Причина подобного явления лежит в достаточно сложных биохимических процессах, связанных с тем, что аллели кодируют несколько разные, но аналогичные по своим функциям полипептиды. В итоге признак как бы накапливается. Сверхдоминирование лежит в основе такого явления как гетерозис, когда дочерние организмы более жизнеспособны, чем родительские.
Помимо перечисленных существуют и другие более редкие типы взаимодействия аллелей.
Глава 4. ОРГАНИЗМЕННЫЙ УРОВЕНЬ ОРГАНИЗАЦИИ ЖИВОГО
4.4. ЗАКОНОМЕРНОСТИ НАСЛЕДОВАНИЯ
4.4.4. Множественный аллелизм
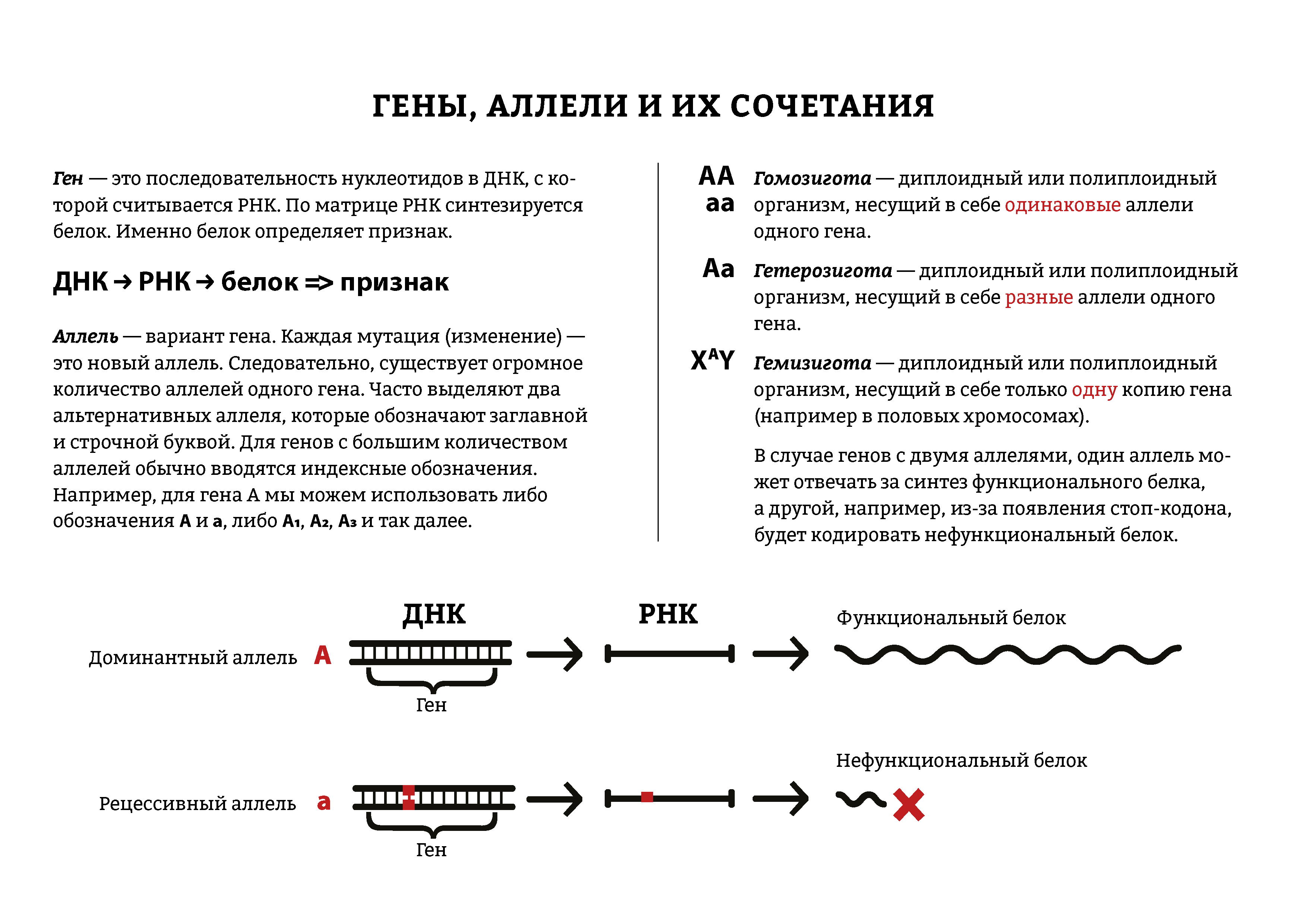
Многие гены у разных организмов существуют более чем в двух аллельных состояниях. Они возникают вследствие мутаций, замены или утраты нуклеотида в молекуле ДНК.
В генотипе диплоидных организмов могут находиться только два гена из серии аллелей. Остальные аллели данного гена в разных сочетаниях входят в генотипы других особей данного вида. Множественные аллели характеризуют разнообразие генофонда вида. Это видовой признак, а не индивидуальный.
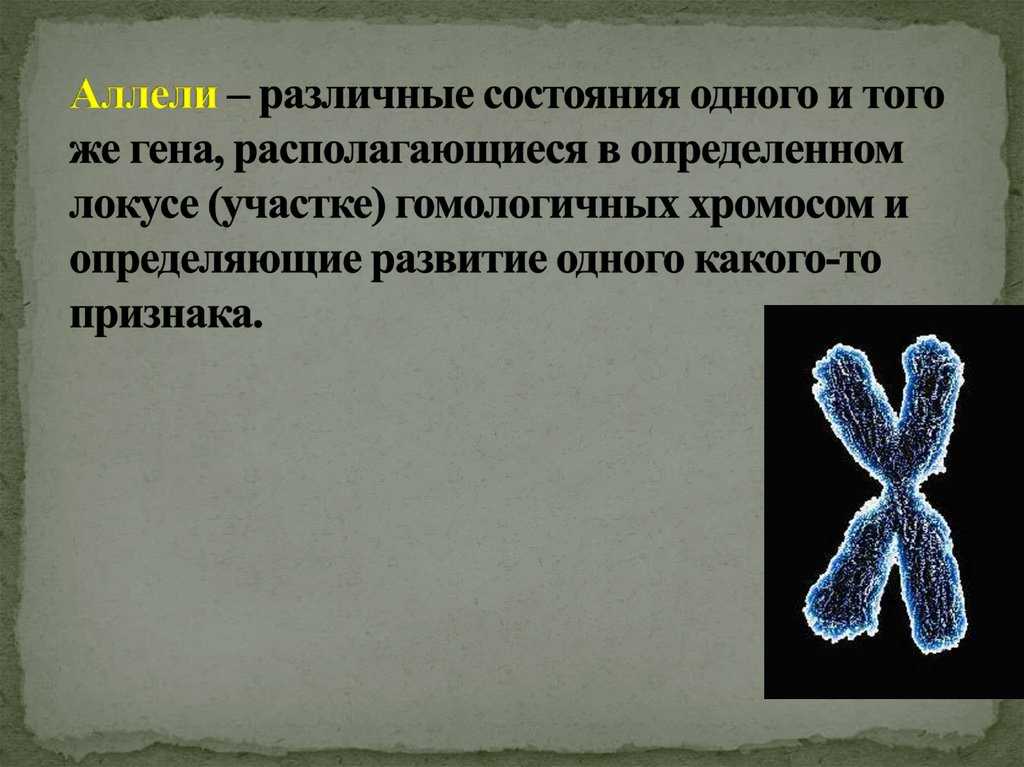
Все аллельные состояния одного и того же гена обозначают одной буквой с разными индексами (рис. 141).
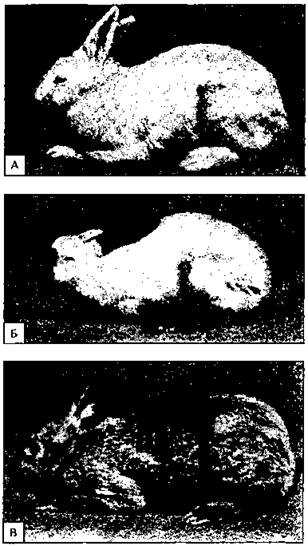
Рис. 141. Множественные аллели у кроликов.
A-альбинос, Б — гималайский, В — темная окраска.
Примером множественных аллелей у человека могут быть гены, отвечающие за развитие АВ0 групп крови: IА, IВ, I. Гены IА и IВ доминантны по отношению к гену J. Проявление действия обоих аллельных генов при одновременном их присутствии называют кодоминантностью. Например, IV группа крови IАIВ проявляется при взаимодействии генов IA и IB. II определяют I группу крови, IAIA, IAI — II группу, IBIB, IBI — III группу.
Группы крови — это иммунологические признаки крови, обусловленные определенными специфическими антигенами, находящимися в эритроцитах, лейкоцитах, тромбоцитах, плазме крови, в тканях и биологических жидкостях человека. Система АВ0 у человека включает 4 основные группы.
I группа не содержит антигенов в эритроцитах, но есть (α- и β-антитела в плазме крови.
II группа в эритроцитах имеет антиген А, а в плазме крови антитела β.
III группа — антиген В, а антитела α.
IV группа имеет антигены А и В, а антител в плазме нет.
При взаимодействии одноименных антигенов и антител происходит агглютинация эритроцитов. Этим обусловлена несовместимость по группам крови. Идеально совместима для реципиента кровь той же самой группы. Кровь людей I группы универсальна для всех групп, т.к. в ней нет антигенов. Эти люди — универсальные доноры. Люди с IV группой крови могут быть универсальными реципиентами, т.к. им возможно переливание крови любой группы (табл. 9).
Таблица 9. Исключение отцовства на основании определения групп крови
|
Группа крови матери |
Группа крови ребенка |
Группы крови и их генотипы, не исключающие отцовства |
Фенотипы по группам крови, исключающие отцовство |
|
I(0) |
I(0) |
I(ii), II(Iai), III(IBi) |
IV(AB) |
|
I(0) |
II(A) |
I(ii), II(Iai), III(IBi) |
IV(AB) |
|
I(0) |
III(B) |
I(ii), II(Iai), III(IBi) |
IV(AB) |
|
II(A) |
I(0) |
II(IAIA,IAi), IV(AB) |
I(0), III(B) |
|
II(A) |
II(A) |
I(ii), II(IAi, IAIA), III(IBi), IV(AB) |
— |
|
II(A) |
III(B) |
II(IAi, IAIA), IV(AB) |
I(0), III(B) |
|
II(A) |
IV(AB) |
I(ii), II(IBi, IAIA), III(IBi), IV(IAIB) |
— |
|
III(B) |
I(0) |
III(IBi, IBIB), IV(AB) |
I(0), II(A) |
|
III(B) |
II(A) |
III(IBi, IBIB), IV(AB) |
I(0), II(A) |
|
III(B) |
III(B) |
I(ii), II(Iai), III(IBi, IBIB), IV(IAIB) |
— |
|
III(B) |
IV(AB) |
I(ii), II(Iai), III(IBi, IAIB), IV(IAIB) |
— |
|
IV(AB) |
II(A) |
III(IBi, IBIB), IV(IAIB) |
I(0), II(A) |
|
IV(AB) |
III(B) |
II(IAi, IAIA), IV(IAIB) |
I(0), III(B) |
|
IV(AB) |
IV(AB) |
II(IAi, IAIA), III(IBi, IBIB), IV(AB) |
I(0) |
Каждому антигену соответствует определенный ген, каждой группе крови — свой генотип.
|
Группа крови |
Ген |
Генотип |
|
I группа |
I |
II |
|
II группа |
IA |
IAIA IAI |
|
III группа |
IB |
IBIB, IBI |
|
IV группа |
IAIB |
IAIB |
Наследование групп крови происходит по законам Менделя.
ПредыдущаяСледующая
Cookie файлы бывают различных типов:
Необходимые. Эти файлы нужны для обеспечения правильной работы сайта, использования его функций. Отключение использования таких файлов приведет к падению производительности сайта, невозможности использовать его компоненты и сервисы.




Файлы cookie, относящиеся к производительности, эффективности и аналитике. Данные файлы позволяют анализировать взаимодействие посетителей с сайтом, оптимизировать содержание сайта, измерять эффективность рекламных кампаний, предоставляя информацию о количестве посетителей сайта, времени его использования, возникающих ошибках.
Рекламные файлы cookie определяют, какие сайты Вы посещали и как часто, какие ссылки Вы выбирали, что позволяет показывать Вам рекламные объявления, которые заинтересуют именно Вас.
Электронная почта. Мы также можем использовать технологии, позволяющие отслеживать, открывали ли вы, прочитали или переадресовывали определенные сообщения, отправленные нами на вашу электронную почту. Это необходимо, чтобы сделать наши средства коммуникации более полезными для пользователя. Если вы не желаете, чтобы мы получали сведения об этом, вам нужно аннулировать подписку посредством ссылки «Отписаться» («Unsubscribe»), находящейся внизу соответствующей электронной рассылки.
Сторонние веб-сервисы. Иногда на данном сайте мы используем сторонние веб-сервисы. Например, для отображения тех или иных элементов (изображения, видео, презентации и т. п.), организации опросов и т. п. Как и в случае с кнопками доступа к социальным сетям, мы не можем препятствовать сбору этими сайтами или внешними доменами информации о том, как вы используете содержание сайта.
Частота аллелей
Частота встречаемости аллельных генов в популяции может быть различной. Обычно рецессивные гены редки и по-сути являются мутациями доминантного аллеля. Многие мутации являются вредными. Однако именно мутантные гены создают материал для действия естественного отбора и как следствие процесса эволюции.
В гипотетической идеальной популяции (в которой не действует естественный отбор, которая имеет неограниченно большой размер, изолирована от других популяций и так далее) частота генотипов (по тому или иному гену) не меняется и подчиняется закону Харди-Вайнберга. Согласно этому закону распределение генотипов в популяции будет укладываться в уравнение: p2 + 2pq + q2 = 1. Здесь p и q — частоты (выраженные в долях единицы) аллелей в популяции, p2 и q2 — частоты соответствующих гомозигот, а 2pq — частота гетерозигот.