

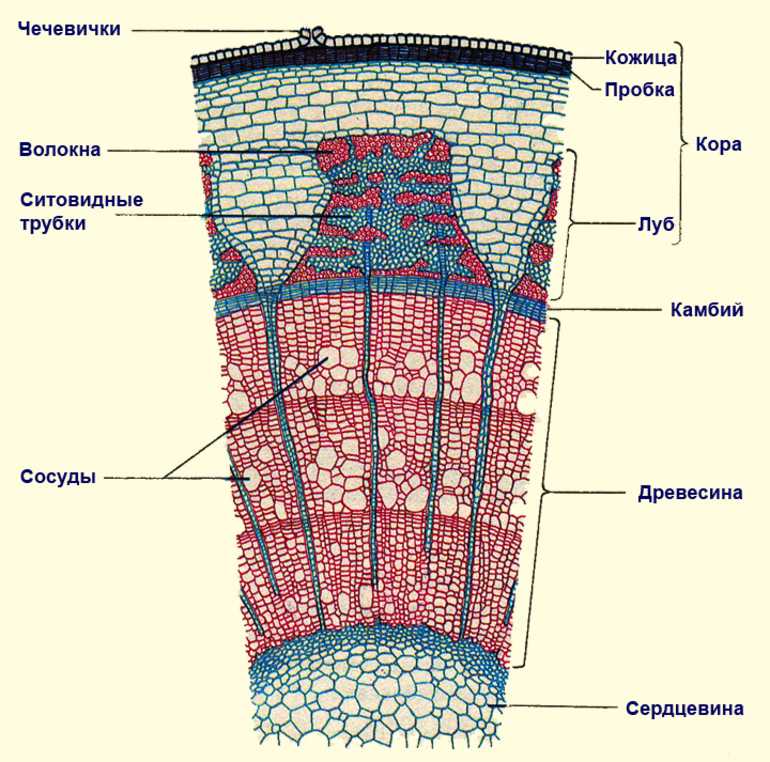
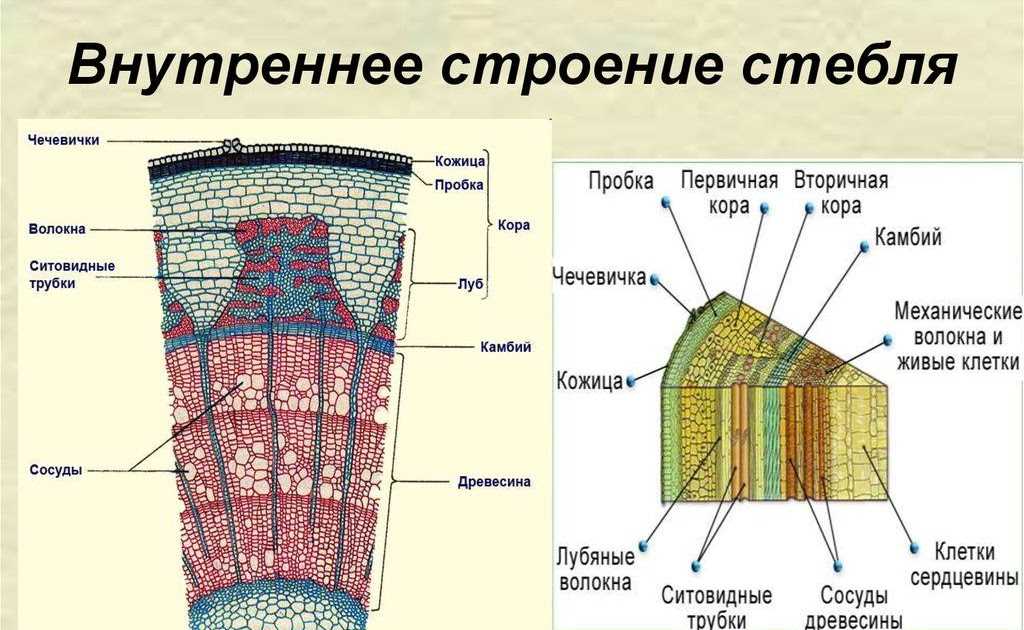
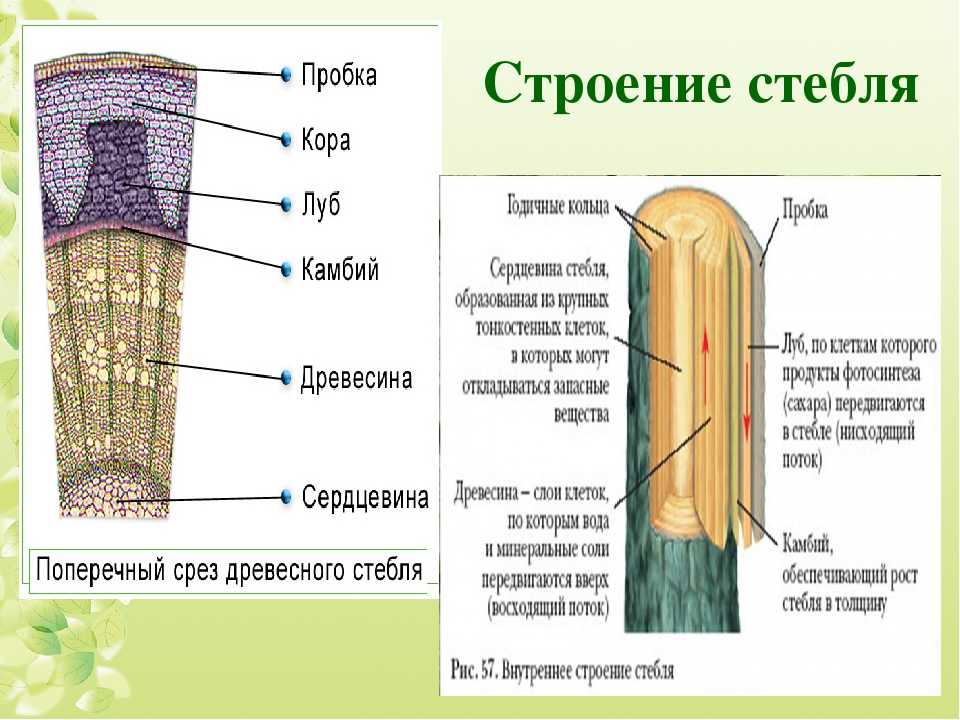
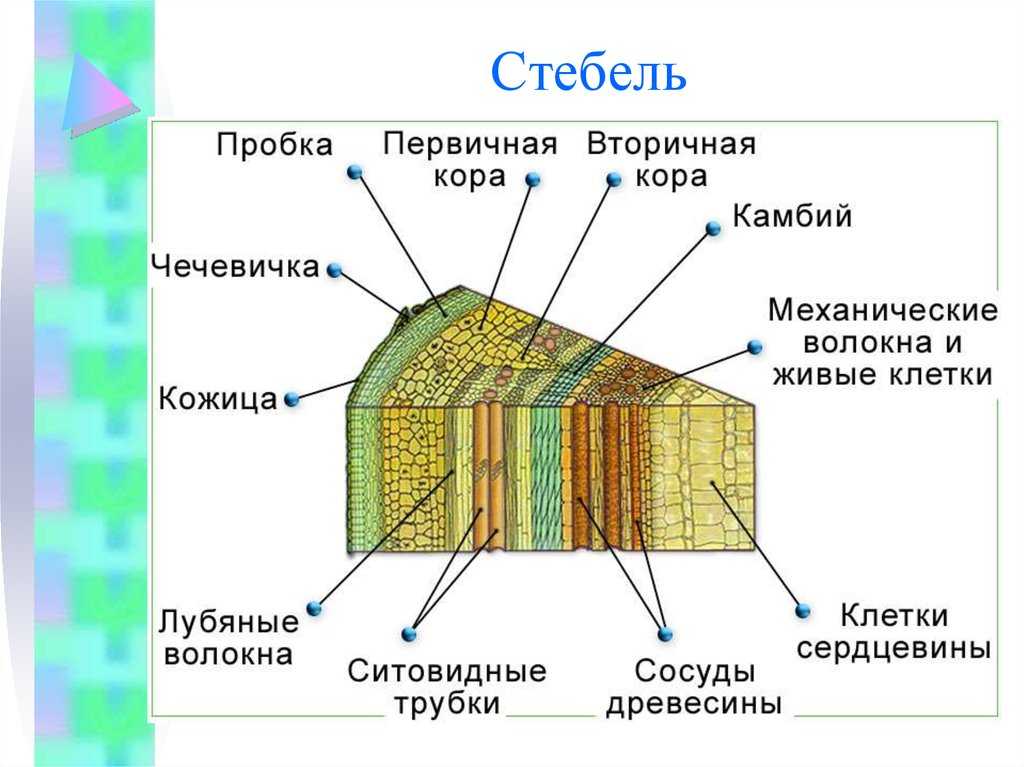
Анатомия стебля
Основные функции стебля — проведение воды и питательных веществ. Дополнительными функциями могут быть фотосинтез, обычно происходящий в молодых стеблях растений, и отложение запасных питательных веществ в многолетних стеблях.
Анатомическое строение стебля растений обусловлено его главными функциями.
Для него характерно развитие механической и проводящей тканей растений. Кроме того, для стебля характерна сложная система меристем — верхушечные, боковые и вставочные, определяющие нарастание его в течение длительного времени и возникновение новых органов.
Стебель растения возникает из апикальной меристемы, из которой дифференцируются три слоя тканей: покровная, проводящая, основная.
Сначала эти ткани представлены так называемой первичной меристемой (промеристемой), состоящей из протодермы, прокамбия и основной меристемы.
Первичные проводящие ткани развиваются из прокамбия.
Первые элементы флоэмы дифференцируются из наружных, расположенных к периферии клеток прокамбия. Первичная флоэма представлена тонкостенными недолговечными удлиненными клетками и носит название протофлоэмы, а наружные клетки ее могут быть представлены механическими волокнами.
Первичные элементы ксилемы — трахеиды, реже сосуды с кольчатыми и спиральными утолщениями стенок — возникают позже из внутренних клеток прокамбия и определяются в целом как протоксилема.
В ее состав помимо проводящих элементов входят паренхимные клетки.
Позже внутрь от протофлоэмы дифференцируется имеющая более или менее типичное для флоэмы строение. Наружу от протоксилемы формируется метаксилема, состоящая из трахеид или трахей с более утолщенными одревесневшими стенками.
Таким образом, за счет деятельности прокамбия и остальной меристемы апекса возникает первичное строение стебля растения.
У однодольных растений весь прокамбий дифференцируется в элементы первичных проводящих тканей.
Механические элементы
Механические элементы луба представлены лубяными волокнами и каменистыми клетками. Первые, соединяясь группами, образуют длинные тяжи, окутывающие ствол в виде сетки. Наряду с настоящими лубяными волокнами иногда встречаются, подобно перегородчатому либриформу древесины, перегородчатые лубяные волокна, разделенные тонкими перегородками. Кроме лубяных волокон механическую роль играют каменистые клетки, которые образуются из паренхимы путем утолщения оболочек. Последние при этом всегда древеснеют и клетки отмирают.
Механические ткани — не необходимый элемент луба. У сосны, например, он совершенно отсутствует, у других древесных пород имеются только лубяные волокна (липа) или только каменистые клетки (ель, береза), и наконец у третьей категории те и другие (дуб, ива, ольха, клен).
Расположение тканей луба не имеет той правильности и периодичности, которые наблюдаются в древесине. Можно только наблюдать чередование слоев тонкостенных элементов (ситовидных трубок и паренхимы) с толстостенными лубяными волокнами. В этом случае говорят о толстостенном и тонкостенном лубе. У липы, дуба и можжевельника чередование того и другого луба довольно правильно и хорошо заметно, причем ежегодно образуется 2 слоя толстостенного луба.
Строение
Рис. 1. Главные разрезы ствола дерева: П – поперечный; Р – радиальный; Т – тангенциальный.
Рис. 2. Схема микроскопического строения древесины сосны (по В. Е. Вихрову): 1 – годичный слой; 2 – многорядный луч с горизонтальным смоляным ходом; 3 – окаймлённая пора; 4 – л…
Д. изучают на трёх разрезах ствола: поперечном и двух продольных – радиальном и тангенциальном (рис. 1). В Д. различают заболонь (периферич. светлую зону) и ядро (центр. зону), имеющее более тёмную окраску у т. н. ядровой Д. или мало отличающееся по цвету от заболони у безъядровой Д. Среди безъядровых пород (ель, пихта, бук и др.) выделяют спелодревесные, у которых центр. зона Д. в свежесрубленном состоянии менее влажная, чем периферическая, и заболонные (берёза, клён) – с равномерной влажностью по сечению ствола. Годичные слои (ежегодные приросты Д.) на поперечном разрезе имеют вид концентрич. окружностей, на радиальном и тангенциальном – соответственно прямых и изогнутых полос; у мн. пород в каждом слое заметны менее плотная светлая (т. н. ранняя) и более плотная тёмная (поздняя) Д. У кольцесосудистых лиственных пород (напр., дуб, ясень) крупные сосуды расположены только в ранней Д., а у рассеянно-сосудистых (берёза, осина) крупные и мелкие сосуды равномерно распределены по годичному слою. У некоторых лиственных пород на поперечном разрезе видны светлые радиальные полоски (лучи), на радиальном – блестящие тёмные или светлые поперечные полоски, а на тангенциальном – веретеновидные узкие полоски. У некоторых хвойных пород (сосна, кедр и др.) в поздней зоне годичных слоёв на поперечном разрезе заметны светлые пятнышки – смоляные ходы.
Рис. 3. Схема микроскопического строения древесины берёзы (по В. Е. Вихрову): 1 – волокнистые трахеиды; 2 – годичный слой; 3 – лучи; 4 – сосуды.
Наблюдаемая с помощью оптич. и электронных микроскопов структура Д. срубленного дерева включает растительные клетки с отмершим протопластом (т. н. мезоструктура). Клеточные стенки (микроструктура) состоят в осн. из целлюлозных микрофибрилл (наноструктура). В тонкой первичной и толстой трёхслойной вторичной оболочке клеточной стенки микрофибриллы имеют разл. ориентацию; в наиболее мощном внутр. слое вторичной оболочки микрофибриллы расположены под небольшим углом наклона (5–15°) к длинной оси клетки. Такая преимущественная ориентация микрофибрилл – одна из осн. причин анизотропии Д. Со стороны полости клетки стенку покрывает тонкий бородавчатый слой. В стенках клеток имеются простые или окаймлённые поры. В промежутках между микрофибриллами находится лигнин, вызывающий одревеснение клеточных стенок, а также гемицеллюлозы и вода.
Д. хвойных пород в осн. состоит из удлинённых прозенхимных клеток – трахеид (рис. 2). Расположенные в ранней зоне годичного слоя крупнополостные трахеиды выполняют гл. обр. проводящую функцию, поздние толстостенные трахеиды – механическую, а паренхимные клетки, образующие лучи и участвующие в структуре вертикальных смоляных ходов, – запасающую. Горизонтальные ходы в некоторых лучах пересекаются с вертикальными, составляя единую смолоносную систему. В Д. лиственных пород (рис. 3) проводящую функцию выполняют сосуды, сосудистые и волокнистые трахеиды; механическую – волокна либриформа и/или волокнистые трахеиды; запасающую – паренхимные клетки в виде горизонтальных однорядных и многорядных лучей, а также вертикальной осевой паренхимы.
Строение почек
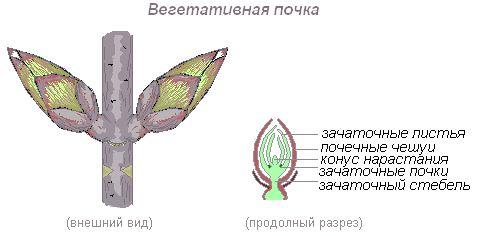
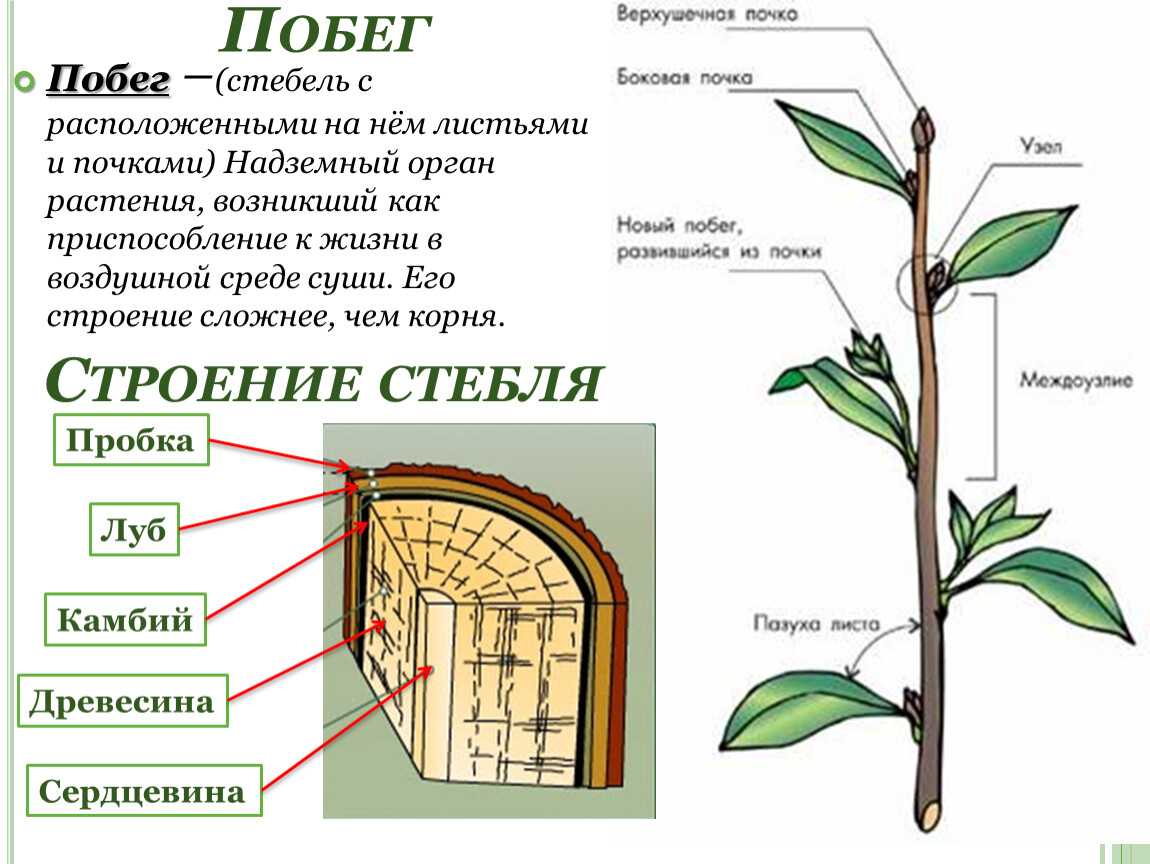
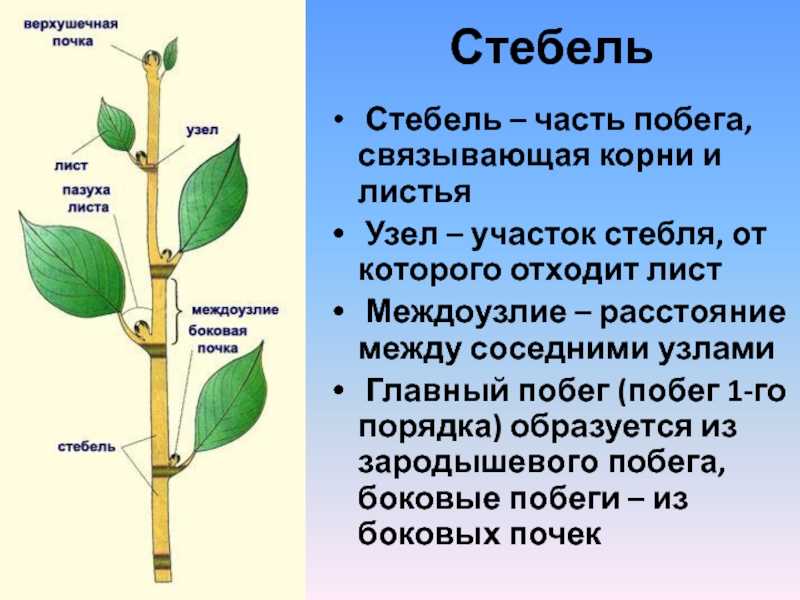
Почка — зачаточный, ещё не развернувшийся побег, на верхушке которого находится конус нарастания.
Вегетативная (листовая почка) — почка, состоящая из укороченного стебля с зачаточными листьями и конуса нарастания.
Генеративная (цветочная) почка — почка, представленная укороченным стеблем с зачатками цветка или соцветия. Цветочная почка, заключающая 1 цветок, называется бутоном.






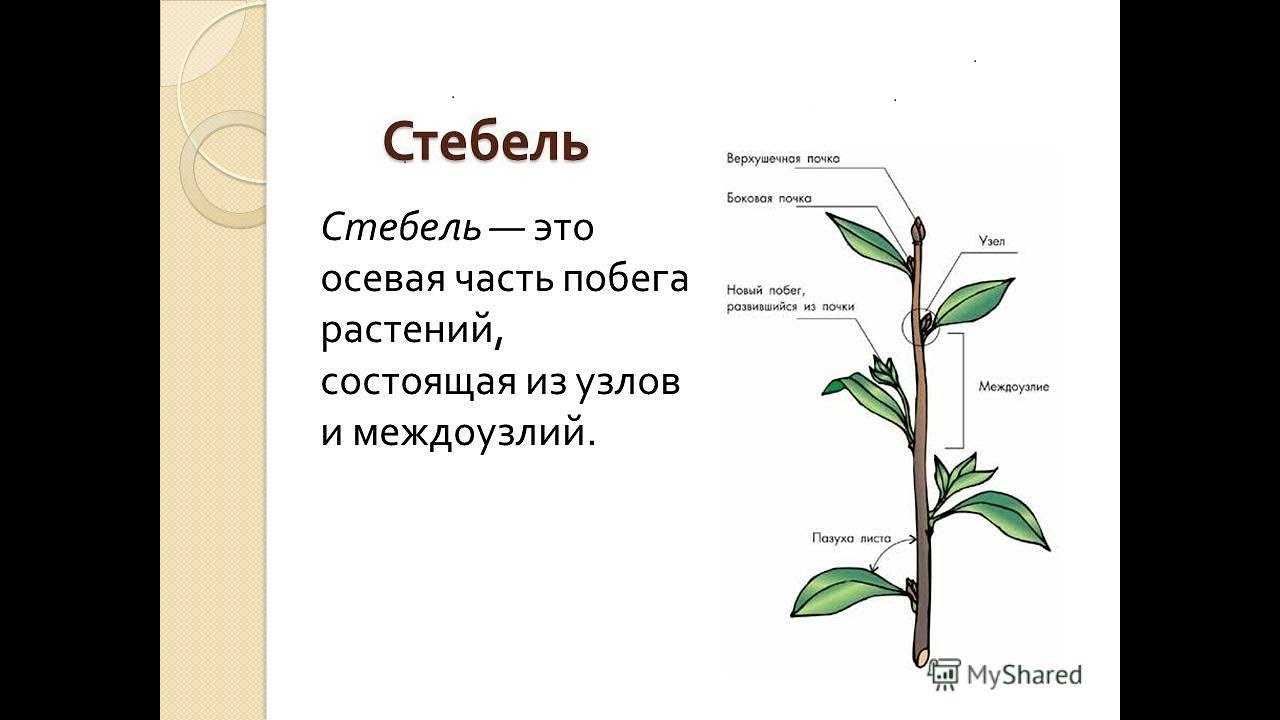
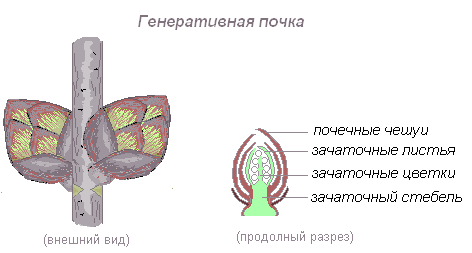
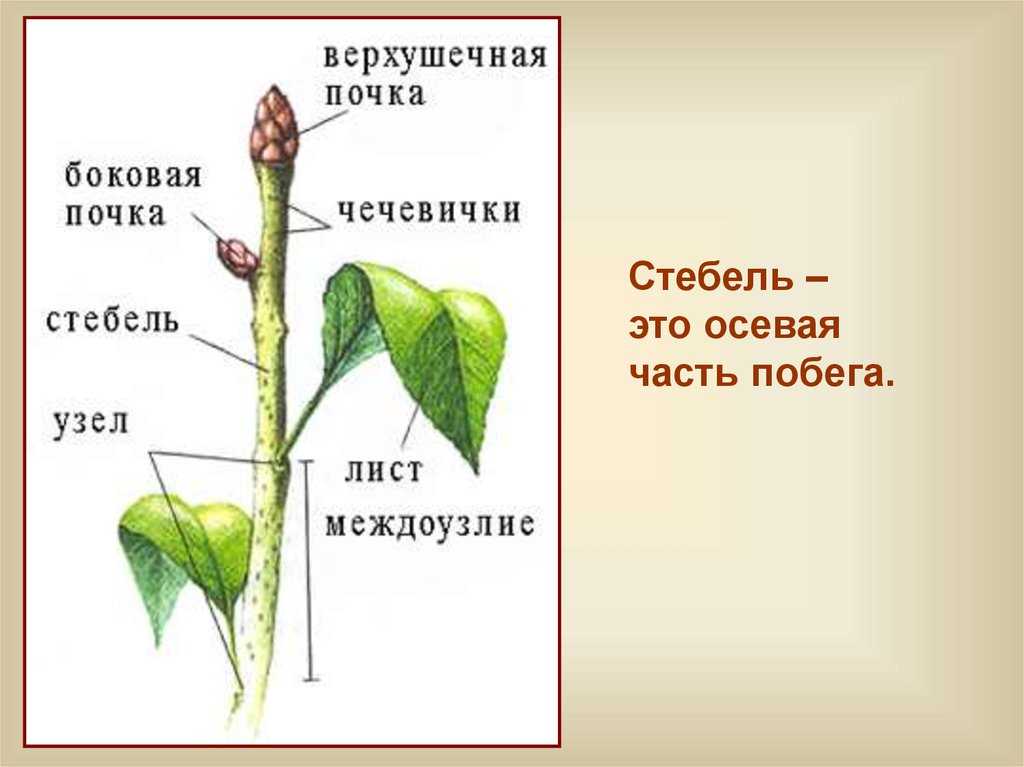
Верхушечная почка — почка, расположенная на верхушке стебля, прикрытая молодыми зачатками листьев, налегающими друг на друга. За счёт верхушечной почки побег нарастает в длину. Она оказывает тормозящее действие на пазушные почки; удаление её приводит к активности спящих почек. Тормозные реакции нарушаются, и почки распускаются.
На верхушке зачаточного стебля находится ростовая часть побега — конус нарастания. Это верхушечная часть стебля или корня, состоящая из образовательной ткани, клетки которой постоянно делятся путём митоза и дают прирост органу в длину. На верхушке стебля конус нарастания защищён почечными чешуевидными листьями, в нём закладываются все элементы побега — стебель, листья, почки, соцветия, цветки. Конус нарастания корня защищён корневым чехликом.
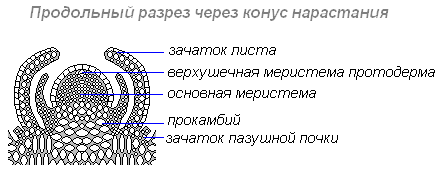
Боковая пазушная почка — почка, возникающая в пазухе листа, из которой образуется боковой побег ветвления. Пазушные почки имеют такое же строение, как и верхушечная. Боковые ветви, следовательно, также растут своими верхушками, и на каждой боковой ветви конечная почка также является верхушечной.
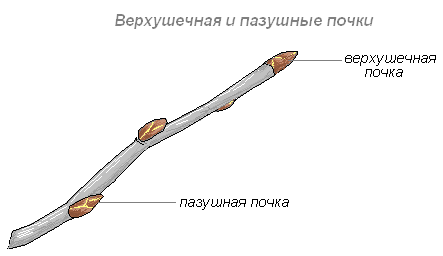
На вершине побега обычно располагается верхушечная почка, а в пазухах листьев — пазушные почки.
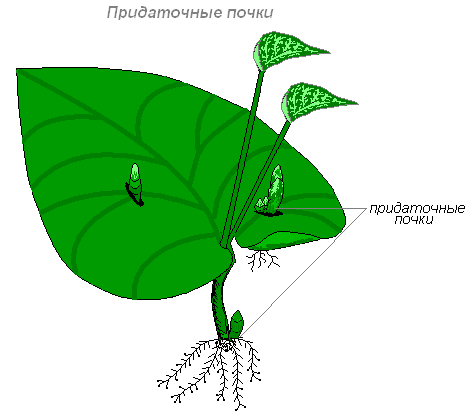
Кроме верхушечных и пазушных почек, у растений часто образуются так называемые придаточные почки. Эти почки не имеют определенной правильности в расположении и возникают из внутренних тканей. Источником их образования могут быть перицикл, камбий, паренхима сердцевинных лучей. Придаточные почки могут образовываться и на стеблях, и на листьях, и даже на корнях. Однако по строению эти почки ничем не отличаются от обычных верхушечных и пазушных. Они обеспечивают интенсивное вегетативное возобновление и размножение и имеют большое биологическое значение. В частности, при помощи придаточных почек размножаются корнеотпрысковые растения.
Спящие почки. Не все почки реализуют свою возможность вырасти в длинный или короткий годичный побег. Некоторые почки не развёртываются в побеги в течение многих лет. При этом они остаются живыми, способными при определённых условиях развиться в листовой или цветоносный побег.
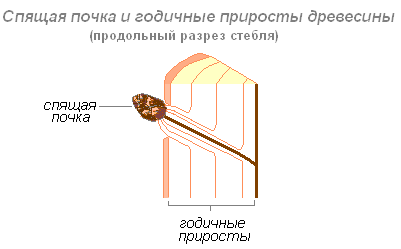
Они словно спят, поэтому их и назвали спящими почками. Когда главный ствол замедляет свой рост или его спиливают, спящие почки трогаются в рост, и из них вырастают облиственные побеги. Таким образом, спящие почки — это очень важный резерв для отрастания побегов. И даже без внешних повреждений старые деревья за счёт них могут «омолаживаться».
Спящие почки, очень характерные для лиственных деревьев, кустарников и ряда многолетних трав. Эти почки не превращаются в нормальные побеги в течение многих лет, нередко они спят в течение всей жизни растения. Обычно спящие почки ежегодно нарастают, ровно настолько, насколько утолщается стебель, именно поэтому они не погребаются нарастающими тканями. Стимулом для пробуждения спящих почек служит обычно гибель ствола. При порубке березы, например, из таких спящих почек образуется пневая поросль. Особую роль спящие почки играют в жизни кустарников. От дерева кустарник отличается своей многоствольностью. Обычно у кустарников главный материнский стволик функционирует недолго несколько лет. При затухании роста главного стволика пробуждаются спящие почки и из них формируются дочерние стволики, которые в росте обгоняют материнский. Таким образом, сама кустарниковая форма возникает в результате деятельности спящих почек.
Смешанная почка — почка, состоящая из укороченного стебля, зачаточных листьев и цветков.
Почка возобновления — зимующая почка многолетнего растения, из которой развивается побег.
Паренхима луба
Паренхима луба, как и паренхима древесины, состоит из паренхимы сердцевинных лучей и паренхимных волокон. От паренхимы древесины она отличается целлюлозными оболочками и более разнообразным содержимым. В ней часто встречаются не только крахмал и масло, но и кристаллы щавелевокислого кальция (одиночные и в сростках — друзах), дубильные вещества и различные сильнодействующие, ценные в медицине вещества, например салицин, хинин и др. Благодаря этому кора некоторых древесных пород, как например дуба, ели, ивы применяется для дубления, а кора крушины, хинного дерева и др. в медицине. В растении эти вещества играют некоторую защитную роль, предохраняя их от повреждений разными паразитами, а также от огладывания зайцами, мышами, козами и т. д.
Паренхимные клетки с кристаллами иногда располагаются в продольном направлении с большой правильностью, прилегая к лубяным волокнам. Тогда они носят особое название кристаллоносной паренхимы и играют большую роль при определении древесных пород по строению коры. Подобно заменяющим волокнам древесины, в лубе также встречаются живые клетки в виде волокон, не разделенных поперечными перегородками на плренхиматические клетки. Эти нежные живые клетки мало отличаются по форме от произведших их клеток камбия и поэтому носят название камбиформа. Паренхимные клетки иногда, как например у сосны, лиственницы и других хвойных, располагаются правильными тангентальными полосками, чередующимися с рядами ситовидных трубок. Ежегодно образуется одна, реже две полоски.
Паренхимная клетка
Паренхимные клетки образуют сердцевинные лучи, а также ряды вертикальной паренхимы. Сердцевинные лучи в древесине лиственных пород развиты сильнее ( от 5 до 34 % объема ствола) и крупнее по размеру, чем у хвойных пород. Вертикальная паренхима либо окружает сосуды, непосредственно контактируя с ними, либо располагается в ксилеме независимо от сосудов.
Паренхимные клетки мезофилла содержат специализированные органеллы — хлоропласта, — осуществляющие фотосинтез. В хлоропластах находится хлорофилл. У двудольных клетки палисадной паренхимы мезофилла, содержащие больше хлоропластов, располагаются вблизи верхней поверхности листа, что обеспечивает максимальное улавливание света. Сравнительно большая длина этих клеток увеличивает возможности поглощения света. Хлоропласты находятся по периферии клеток палисадной паренхимы. Это позволяет им поглощать максимально возможное количество света и облегчает газообмен. У двудольных губчатая паренхима мезофилла имеет обширные межклетники для эффективного газообмена.
|
Схема строения хвойной древесины. |
Часть паренхимных клеток образует радиально расположенные сердцевинные лучи ствола. Эти клетки являются местом, где происходит образование резервных веществ ( крахмала и жиров) осенью и расходование их весной.
|
Схема строения хвойной древесины. |
Часть паренхимных клеток образует радиалыю расположенные сердцевинные лучи ствола. Эти клетки являются местом, где происходит образование резервных веществ ( крахмала и жиров) осенью и расходование их весной.
|
Строение паренхимных клеток. А. Поперечный срез. Клетки обычно изодиаметрические ( округлые, но могут быть и вытянутыми. Б. Поперечный срез сердцевины стебля Helianthus. Сердцевина — это выполняющая и опорная ткань, находящаяся в центре стебля двудольных. |
Стенки паренхимных клеток — важный путь, по которому перемещаются в растении вода и минеральные соли ( часть апопластного пути, который будет описан в гл. Вещества могут перемещаться также и по плазмодесмам, связывающим соседние клетки.
|
Строение эндодермы корня. А. Поперечный срез. Видна молодая эндодерма с поясками Каспари. Б. Поперечный срез старого корня двудольного растения. |
Так называются специализированные паренхимные клетки, примыкающие к ситовидным трубкам и участвующие в их работе. Метаболически клетки-спутницы весьма активны; от обычных паренхимных клеток их отличают более плотная цитоплазма и более мелкие вакуоли. О происхождении, строении и функции клеток-спутниц мы будем говорить в разд.
Кроме того, паренхимные клетки, образуя межклеточные каналы, как бы выстилают поверхность смоляных ходов, столь характерных для хвойных пород.
Третью группу составляют паренхимные клетки, которые образуют основную массу ткани флоэмы. Паренхимные клетки располагаются вертикальными полосами или распределяются среди ситовидных клеток. Флоэма также содержит лучи из паренхимных клеток, подобные по строению лучам ксилемы. Лучи флоэмы продолжают лучи ксилемы и выполняют функцию проведения продуктов метаболизма в радиальном направлении. На некотором расстоянии от камбия радиальное расположение лучей нарушается, они принимают в поперечном сечении волнистый вид.




Стенки сосудов и паренхимных клеток по ориентации микрофибрилл несколько отличаются от стенок волокон. Определенные особенности в ориентации целлюлозных микрофибрилл характерны для клеточных стенок реактивной древесины. В стенках трахеид сжатой древесины в слое Sj угол ориентации близок к 45, т.е. намного больше, чем у нормальной древесины. В стенках волокон тяговой древесины в Q-слое микрофибриллы ориентированы почти параллельно оси волокна. Изучение окаймленных пор показало, что в торусах мембран наблюдается кольцевая ориентация микрофибрилл целлюлозы, а в окружающей торус маргинальной ( краевой) зоне мембраны тяжи микрофибрилл ориентированы радиально и удерживают торус. В заболонной древесине торусы не лигнифицированы; отложение в них лигнина происходит при образовании ядровой или спелой древесины.
|
Трахеиды сосны. |
Живой частью древесины являются паренхимные клетки, заполненные плазмой и продуктами обмена веществ. Паренхимные клетки представляют собой обычно короткую прямоугольную четырехгранную призму. Сосна не имеет древесной паренхимы; сердцевидные лучи сосны и ели занимают 5 — 8 % общего объема древесины.
Видоизменения подземных побегов
Корневище — подземный побег, выполняющий функции отложения запасных веществ, возобновления, а иногда и вегетативного размножения. Корневище не имеет листьев, но имеет хорошо выраженную метамерную структуру, узлы выделяются либо по листовым рубцам и остаткам сухих листьев, либо по листовым рубцам и остаткам сухих листьев, либо по живым чешуевидным листьям и по расположению пазушных почек. На корневище могут формироваться придаточные корни. Из почек корневища вырастают его боковые ответвления и надземные побеги.
Корневища характерны преимущественно для травянистых многолетников — копытень, фиалка, ландыш, пырей, земляника и др., но бывают у кустарников и кустарничков. Длительность жизни корневищ колеблется от двух-трёх до нескольких десятков лет.
Клубни — утолщённые мясистые части стебля, состоящие из одного или нескольких междоузлий. Бывают надземными и подземными.
Надземные — утолщение главного стебля, боковых побегов. Часто имеют листья. Надземные клубни являются вместилищем запасных питательных веществ и служат для вегетативного размножения, на них могут находиться метаморфизированные пазушные почки с зачатками листьев, которые опадают и служат также для вегетативного размножения.
Подземные клубни — утолщение подсемядольного колена или подземных побегов. На подземных клубнях листья редуцированы до чешуек, которые опадают. В пазухах листьев находятся почки — глазки. Подземные клубни обычно развиваются на столонах — дочерних побегах — из почек, расположенных у основания основного побега, выглядят как очень тонкие белые стебельки, несущие маленькие бесцветные чешуевидные листья, растут горизонтально. Клубни развиваются из верхушечных почек столонов.
Луковица — подземный, реже надземный побег с очень коротким утолщённым стеблем (донцем) и чешуевидными мясистыми, сочными листьями, запасающие воду и питательные вещества, в основном сахара. Из верхушечной и пазушных почек луковиц вырастают надземные побеги, а на донце образуются придаточные корни. В зависимости от размещения листьев различают луковицы чешуевидные (лук), черепитчатые (лилия) и сборные или сложные (чеснок). В пазухе некоторых чешуек луковицы есть почки, из которых развиваются дочерних луковицы — детки. Луковицы помогают растению выжить в неблагоприятных условиях и являются органом вегетативного размножения.
Клубнелуковицы — внешне похожи на луковицы, но их листья не служат запасающими органами, они сухие, плёнчатые, часто это остатки влагалищ отмерших зелёных листьев. Запасающий орган — стеблевая часть клубнелуковицы, она утолщена.
Надземные столоны (плети) — недолговечные ползучие побеги, служащие для вегетативного размножения. Встречаются у многих растений (костянка, полевица, земляника). Обычно они лишены развитых зелёных листьев, стебли их тонкие, хрупкие, с очень длинными междоузлиями. Верхушечная почка столона, загибаясь вверх, даёт розетку листьев, которая легко укореняется. После укоренения нового растения столоны разрушаются. Народное название этих надземных столонов — усы.
Колючки — укороченные побеги, имеющие ограниченный рост. У некоторых растений формируются в пазухах листьев и соответствуют боковым побегам (боярышник) или образуются на стволах из спящих почек (гледичия). Характерны для растений жарких и сухих мест произрастания. Выполняют защитную функцию.
Побеги суккулентов — надземные побеги, приспособленные для накопления воды. Обычно с образованием суккулентного побега связана потеря или метаморфоз (превращение в колючки) листьев. Сочный стебель выполняет две функции — ассимиляционную и водозапасающую. Характерны для растений, живущих в условиях длительного недостатка влаги. Стеблевые суккуленты наиболее представлены в семействе кактусовых, молочайных.
* * *
Покровные ткани
Рисунок 1: Эпидерма.
Рисунок 2: Основные типы устьичных аппаратов. 1 – диацитный; 2 –парацитный; 3 –анизоцитный; 4 — аномоцитный.
Вторичная покровная ткань высших растений – это пробка. Пробковый слой обычно образуется на вторично утолщенных стеблях и корнях высших растений. Пробка (она же феллема), образуется в результате работы так называемого пробкового камбия (или феллогена). В феллогене клетки делятся и откладываются наружу, их клеточные стенки утолщаются и суберинизируются (опрбковевают). Суберин – это вещество непроницаемое для воды и воздуха, следовательно, внутреннее содержимое клеток вскоре отмирает. В результате пробковый слой состоит из мертвых клеток и является газо- и водонепроницаемой покровной тканью.
Рисунок 3: Феллема, феллоген, феллодерма.
Образовательные ткани
Массив ткани, в которой происходят клеточные деления в теле высшего растения, следует назвать образовательной тканью или меристемой. Образовательные ткани не являются постоянными. Клетки меристемы недифференцированные и не специализированные, у них тонкие клеточные оболочки. Данные клетки делятся и в дальнейшем преобразуются в ту или иную специализированную ткань.
Рисунок 1: Апикальная меристема стебля.
У высших растений выделяют две вторичные латеральные меристематические ткани – камбий и феллоген. Камбий (или сосудистый камбий) закладывается в проводящих пучках стебля или корня между флоэмой и ксилемой. В результате клеточных делений внутрь откладывается ткань, дифференцирующаяся в ксилему, а наружу – будущая флоэма. За счет работы камбия происходит процесс вторичного утолщения стебля или корня. Соответственно, сформированные камбием проводящие ткани будут называться вторичными – вторичная ксилема и вторичная флоэма. Следует помнить, что при вторичном утолщении камбиальная зона возникает не только внутри проводящих пучков, но и формируется так называемый межпучковый камбий. В результате на поперечном срезе камбий имеет вид общего меристематического кольца.
Рисунок 2: Камбий. 1 – эпидерма; 2 – паренхима; 3 – флоэмные волокна; 4 – флоэма; 5 – пучковый камбий; 6 – ксилема; 7 – межпучковый камбий.
Феллоген (или пробковый камбий) возникает в корнях и стеблях растений при их вторичном утолщении. При утолщении эпидерма и впоследствии первичная кора опадает и отмирает, покровную функцию в данном случае выполняет пробковый слой, формируемый феллогеном. В результате клеточных делений в феллогене, наружу откладываются клетки феллемы (или пробка). Феллоген снизу подстилается слоем клеток – феллодермой. Комплекс из трех данных тканей носит название перидерма.
Рисунок 3: Феллема, феллоген, феллодерма.
Образование годичных колец
Годичные кольца. Рассматривая древесину на поперечном срезе через многолетний стебель дерева, можно увидеть чередующиеся светлые и темные концентрические кольца. Каждая пара колец (светлое и темное) – это слой древесины, образовавшийся на протяжении одного года за счет деятельности камбия. Он называется годичным кольцом.







Весной клетки камбия активно делятся. Так возникают крупные клетки древесины, в частности, сосуды большого диаметра и с тонкими стенками. На поперечном срезе они составляют светлую часть годичного кольца. Летом же камбий формирует значительно более мелкие толстостенные сосуды и клетки механической ткани. Так возникает темный слой. Зимой клетки камбия вообще не делятся. Следующей весной его деятельность возобновляется и начинает формироваться новое годичное кольцо.
Годичные кольца могут «поведать» об условиях жизни растения. Так, их количество соответствует возрасту побега древесного растения. Ширина колец зависит от погодных условий в тот или иной год жизни растения. Больший прирост наблюдается в годы, благоприятные по своим погодным условиям. У старых деревьев активность камбия снижена, поэтому годичные кольца узкие, а прирост побегов незначительный.
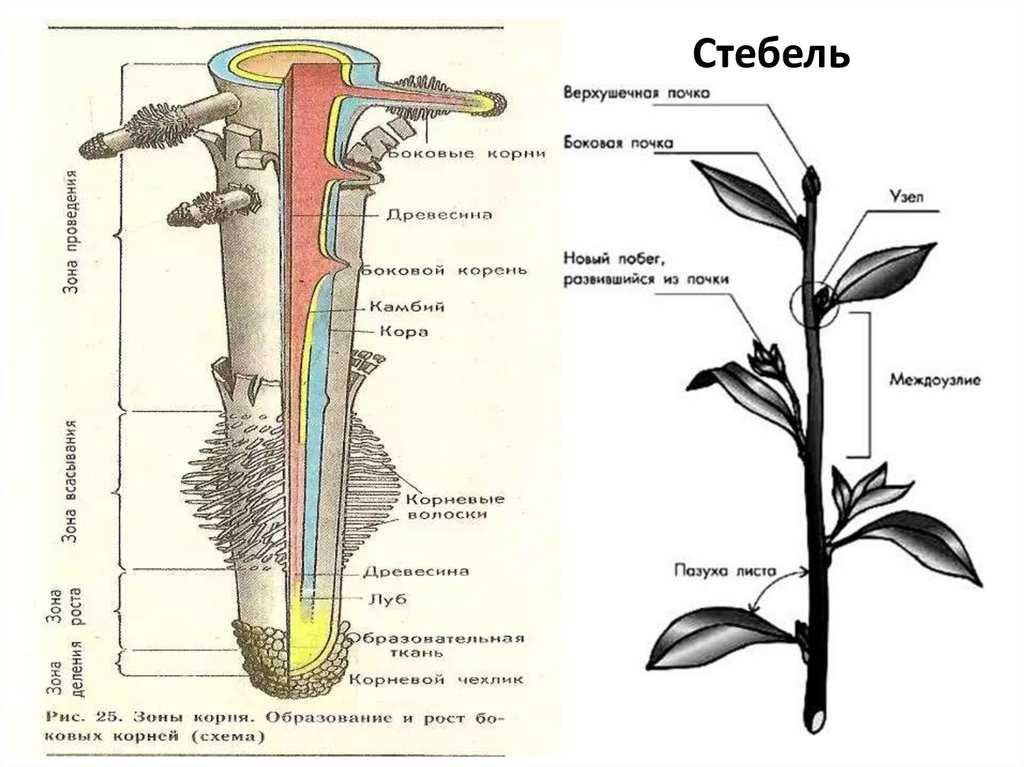
Корень и его основные функции
Корни – это осевые вегетативные органы растений, которые обладают радиальной симметрией.
Чаще всего они располагаются в грунте. На корнях никогда не образуются какие–либо генеративные органы или же листья. Но при этом орган остается одним из основных источников питания.
Подобные вегетативные органы растений служат для поглощения из субстрата воды, и, конечно же, растворенных в ней полезных компонентов.
Помимо этого корень позволяет растению укрепиться в почве. Данный орган синтезирует некоторые органические вещества, например, алкалоиды, гормон роста и другие.
Эти компоненты перемещаются в другие части растения по сосудам ксилемы. Если микроэлементы и гормоны остаются невостребованными, то они сохраняются в корне.
Очень часто эти вегетативные органы растений являются местом для хранения всех питательных компонентов.
Это еще не все функции корня. У некоторых растений, например, у осота полевого, сирени, вишни, малины, ивы, тополя, осины и других этот орган осуществляет вегетативное размножение. У корне отпрысковых на корнях возникают придаточные почки, которые постепенно растут.
Из них в итоге развивается наземная часть.
Вегетативные органы высших растений стали настоящим эволюционным достижением, которое позволило некоторым видам приспособиться к совершенному питанию. Кроме того, корни позволяют образовывать новые побеги, которые могут подыматься вверх, ближе к солнечному теплу и свету.