

Слайд 40 Моногибридное расщепление в F2 у автотетраплоида7. Полиплоидия В
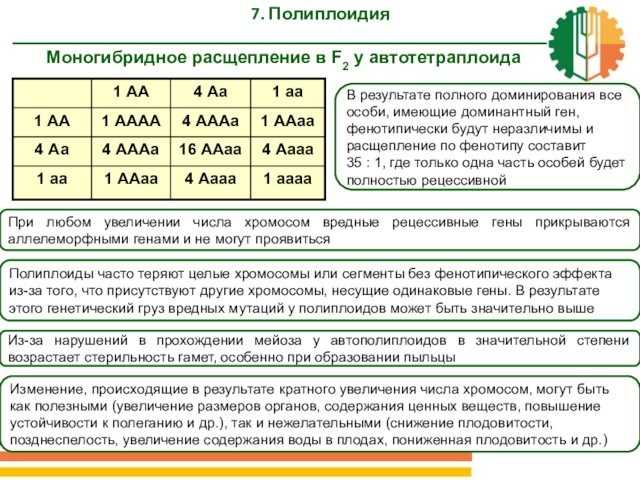
будут неразличимы и расщепление по фенотипу составит 35 : 1,
где только одна часть особей будет полностью рецессивной
При любом увеличении числа хромосом вредные рецессивные гены прикрываются аллелеморфными генами и не могут проявиться
Полиплоиды часто теряют целые хромосомы или сегменты без фенотипического эффекта из-за того, что присутствуют другие хромосомы, несущие одинаковые гены. В результате этого генетический груз вредных мутаций у полиплоидов может быть значительно выше
Из-за нарушений в прохождении мейоза у автополиплоидов в значительной степени возрастает стерильность гамет, особенно при образовании пыльцы
Изменение, происходящие в результате кратного увеличения числа хромосом, могут быть как полезными (увеличение размеров органов, содержания ценных веществ, повышение устойчивости к полеганию и др.), так и нежелательными (снижение плодовитости, позднеспелость, увеличение содержания воды в плодах, пониженная плодовитость и др.)
Слайд 20 И.В. Мичурин для повышения скрещиваемости видов применял следующие

в прививке растений одного вида на растения другого. При сращивании
тканей привитых растений может изменяться химический состав генеративных органов, в результате чего стимулируется прорастание пыльцевых трубок одного вида в тканях пестика другого.
Данный метод используют в случае, когда исходные виды не скрещиваются между собой, но скрещиваются с третьим видом. Получают гибриды с данным видом, а затем скрещивают эти гибриды между собой. Данный метод нашел применение в селекции картофеля, пшеницы, овса и других культур.
Опыление смесью пыльцы. Смесь пыльцы разных видов и разновидностей может способствовать скрещиваемости видов, поскольку пыльцевые трубки с разными генотипами могут взаимно стимулировать рост, создавая в пестике условия, благоприятствующие росту пыльцевых трубок.
4. Причины нескрещиваемости и бесплодия отдаленных гибридов и методы их преодоления
Слайд 27 Восстановление родословной существующих видов экспериментальным путем называется ресинтезом

показана А. Мюнтцингом в начале
30-х годов ХХ века при восстановлении родословной пикульника (Galeopsis). Известно несколько видов этого рода, и среди них G. tetrahit (2n = 32), G. speciosa (2n = 16), G. pubescens (2n = 16).
А. Мюнтцинг скрестил G. speciosa и G. pubescens, но гибриды F1 между ними оказались почти бесплодными. В F2 было лишь одно растение, которое оказалось триплоидом (2n = 24). Это растение повторно опылили пыльцой G. pubescens. Полученные семена дали нормально плодовитые растения, очень похожие на G. tetrahit и имеющие одинаковое с ним число хромосом. Этот искусственный аллополиплоид, легко скрещивающийся с естественным G. tetrahit, был назван G. pseudotetrahit.
Ресинтез проведен в роде пшениц, овса, табака, хлопчатника, сливы, земляники, полиплоидных видов рода капуст и других культур.
Производство
Посевная площадь рапса в мире постоянно увеличивается; его возделывают в Индии, Китае, Канаде и других странах. Основные районы возделывания озимого рапса в СНГ — лесостепная зона Украины, ярового рапса — северная часть лесостепной зоны Украины. Для кормовых целей озимый рапс можно выращивать почти во всех районах степи, лесостепи и лесолуговой зоны России и стран СНГ.
По данным продовольственной и сельскохозяйственной организации ООН в сезоне 2003—2004 годов было собрано 36 млн тонн семян рапса, а в 2004—2005 годах — 46 млн тонн. В 2005 году под рапс было отведено 264 тыс. кв. км, что составляет около 2 % мировой площади пашни. Постоянно растущая рентабельность топливного применения таких культур, как сахарный тростник, рапс, подсолнечник и др., вынуждает сельхозпроизводителей сокращать площади под продовольственными сельскохозяйственными культурами с продовольственными целями. По данным Oil World, мировое производство рапса в 2008—2009 сельскохозяйственном году составило 58 млн тонн. При этом на ЕС пришлось 19 млн тонн, Канаду — 12,6 млн тонн, Китай — 11,5 млн тонн.
Средняя урожайность рапса в мире 15 ц/га (1,5 т/га или 150 т/км²). Максимальная урожайность достигнута белорусским предприятием ОАО «Агрокомбинат «Южный» Гомельского района и составила 64 ц/га (6,4 т/га или 640 т/км²).
Суммарное производство рапса в мире в 2014 году составило 73,8 млн тонн, в 2016 году — 68,9 млн тонн. С 1965 года (5,2 млн тонн) производство выросло в 14 раз, а с 1995 года (34,2 млн тонн) более чем в два раза.
Рапс в России
За последние годы производство рапса в России значительно возросло. В 2006 году посевная площадь под рапсом составила 432 тыс. га, что превысило показатели 2005 года более чем в два раза. На 2009 год динамика роста посевных площадей, занятых под яровой и озимый рапс, была на отметке 694,82 тыс. га. (515,14 тыс. га. — яровой рапс, 179,68 тыс. га. — озимый рапс.) В связи с погодными условиями в основном возделывается яровой рапс. Озимый рапс распространён главным образом на юге, в Ставропольском и Краснодарском краях.
В 2012 году площадь под рапсом в России впервые превысила 1 миллион га, в 2012 году она составила 1 020,7 тыс. га
В России в 2013 году был собран рекордный урожай рапса — 1,39 млн тонн. В 2014 году урожай составил 1,34 млн тонн, в 2015 году — 1,012 млн тонн.
Слайд 23 Нарушение микро- и макроспорогенезаОтсутствие или нарушение коньюгации хромосом

видов. Непосредственными причинами нарушения коньюгации хромосом у отдаленных гибридов F1
являются различные хромосомные аберрации: инверсии, транслокации и т.д., в результате которых у разных видов в процессе эволюции изменяется состав и порядок расположения генов в соответствующих хромосомах и нарушается их парность. Несмотря на парность и частичную гомологичность хромосом, они имеют большие структурные различия.
Коньюгация хромосом у таких гибридов хотя и происходит, но протекает неправильно с образованием большого числа поли- и унивалентов и как следствие этого – неравномерное распределение хромосом между дочерними клетками во время мейоза, образование маложизнеспособных гамет и стерильность большинства гибридов.
4. Причины нескрещиваемости и бесплодия отдаленных гибридов и методы их преодоления
Ссылки[]
| Масличные культуры |
| Арахис | Горчица белая | Горчица сарептская | Горчица чёрная | Клещевина | Кокосовая пальма | Конопля | Кунжут | Лён масличный | Ляллеманция | Маслина | Перилла | Подсолнечник | Рапс | Рыжик | Сафлор | Соя | Тунг | Хлопчатник |
При написании этой статьи использовался материал из Энциклопедического словаря Брокгауза и Ефрона (1890—1907).
Эта страница использует содержимое раздела Википедии на русском языке. Оригинальная статья находится по адресу: Рапс. Список первоначальных авторов статьи можно посмотреть в истории правок. Эта статья так же, как и статья, размещённая в Википедии, доступна на условиях CC-BY-SA .
Учение Н. И. Вавилова о центрах происхождения и многообразия культурных растений
|
Н.И.Вавилов |
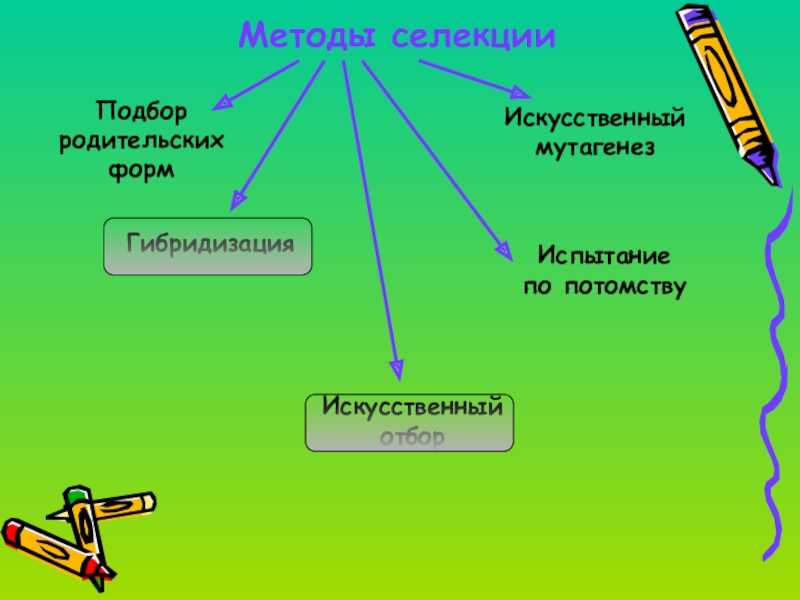
Селекция — наука о создании новых и улучшении существующих пород животных, сортов растений, штаммов микроорганизмов. В основе селекции лежат такие методы, как гибридизация и отбор. Теоретической основой селекции является генетика.
Для успешного решения задач, стоящих перед селекцией, академик Н.И.Вавилов особо выделял значение:
-
Изучения сортового, видового и родового разнообразия интересующей нас культуры;
-
Влияния среды на развитие интересующих селекционера признаков;
-
Изучения наследственной изменчивости;
-
Знаний закономерностей наследования признаков при гибридизации;
-
Особенностей селекционного процесса для само- или перекрестноопылителей;
-
Стратегии искусственного отбора.
Породы, сорта, штаммы — искусственно созданные человеком популяции организмов с наследственно закрепленными особенностями: продуктивностью, морфологическими, физиологическими признаками.
Каждая порода животных, сорт растений, штамм микроорганизмов приспособлены к определенным условиям, поэтому в каждой зоне нашей страны имеются специализированные сортоиспытательные станции и племенные хозяйства для сравнения и проверки новых сортов и пород.
Для успешной работы селекционеру необходимо сортовое разнообразие исходного материала, с этой целью Н.И.Вавиловым была собрана коллекция сортов культурных растений и их диких предков со всего земного шара. К 1940 году во Всесоюзном институте растениеводства насчитывалось 300 тыс. образцов. Но с позиций лысенковщины, занявшей в то время руководящие позиции в биологической науке России и считавшей, что определяющую роль в создании новых форм играет окружающая среда, эта коллекция была не нужна. Работы по пополнению коллекции были прекращены. В настоящее время коллекция пополняется и является основой для работ по селекции любой культуры.









Н.И.Вавилов установил центры происхождения культурных растений, где находится наибольшее видовое и сортовое многообразие культурных растений.
Центры происхождения культурных растений (по Н.И.Вавилову).
|
Центры происхождения |
Местоположение |
Культивируемые растения |
|
1. Южноазиатский тропический 2. Восточноазиатский 3. Юго-Западноазиат-ский 4. Средиземноморский 5. Абиссинский 6. Центральноамериканский 7. Южноамериканский |
Тропическая Индия, Индокитай, о-ва Юго-Восточной Азии Центральный и Восточный Китай, Япония, Корея, Тайвань Малая Азия, Средняя Азия, Иран, Афганистан, Юго-Западная Индия Страны по берегам Средиземного моря Абиссинское нагорье Африки Южная Мексика Западное побережье Южной Америки |
Рис, сахарный тростник, цитрусовые, баклажаны и др. (50% культурных растений) Соя, просо, гречиха, плодовые и овощные культуры — слива, вишня и др. (20% культурных растений) Пшеница, рожь, бобовые культуры, лен, конопля, репа, чеснок, виноград и др. (14% культурных растений) Капуста, сахарная свекла, маслины, клевер (11% культурных растений) Твердая пшеница, ячмень, кофейное дерево, бананы, сорго Кукуруза, какао, тыква, табак, хлопчатник Картофель, ананас, хинное дерево. |
Наиболее богатыми по количеству культур являются древние центры цивилизации, именно там наиболее ранняя культура земледелия, более длительное время проводится искусственный отбор и селекция растений.
Этапы селекции растений
Методы селекции растений претерпели множество изменений с тех пор, как они были начаты 9000-1000 лет назад. Современный метод селекции растений осуществляется в следующие этапы:
- Изменчивость лежит в основе всех методов разведения. Первый шаг включает в себя сбор растений или семян для всех возможных аллелей для всех генов в данной культуре, которая известна как зародышевая плазма. В эту коллекцию входят даже дикорастущие сорта и родственники культивируемых видов.
- Оценка и отбор родительских растений. Зародышевая плазма оценивается для отбора родительских растений с желаемыми характеристиками. Сочетание этих характеристик ожидается в гибридном потомстве. Например, растительная культура с высоким содержанием белка может быть выбрана для скрещивания с растением с более высокой устойчивостью к болезням.
- Перекрестная гибридизация между выбранными родителями. На третьем этапе родители подвергаются перекрестной гибридизации для получения чистых линий потомства. Это утомительная и трудоемкая практика, осуществляемая обычным способом внесения пыльцы с одного растения на рыльце другого. Несмотря на затраченный труд, только одно или два потомства из нескольких сотен демонстрируют желаемое сочетание характеристик.
- Отбор и тестирование превосходных рекомбинантов. Затем оценивают развитое потомство, и те, которые обладают желаемой комбинацией характеристик, самоопыляются для достижения гомозиготности.
- Тестирование, выпуск и коммерциализация новых сортов. Новые сорта выращиваются на исследовательских полях, где они тестируются на их агрономические характеристики качества, урожайности, устойчивости к болезням и т.д. За этим следует выращивание этих культур на полях фермеров в разных местах страны, которые представляют различные агроклиматические зоны. При успешных результатах посевы выпускаются в коммерческих целях для общественного потребления.
Отбор
В селекции действует естественный и искусственный отбор. Искусственный отбор бывает бессознательный и методический. Бессознательный отбор проявляется в сохранении человеком лучших особей для разведения и употреблении в пищу худших без сознательного намерения вывести более совершенный сорт или породу. Методический отбор осознанно направлен на выведение нового сорта или породы с желаемыми качествами.
В процессе селекции наряду с искусственным отбором не прекращает своего действия и естественный отбор, который повышает приспособляемость организмов к условиям окружающей среды.
| Признак | Естественный отбор | Искусственный отбор |
|---|---|---|
| Исходный материал для отбора | Индивидуальные признаки организмов | Индивидуальные признаки организмов |
| Отбирающий фактор | Условия среды (живая и неживая природа) | Человек |
| Путь благоприятных изменений | Остаются, накапливаются, передаются по наследству | Отбираются, становятся производительными |
| Путь неблагоприятных изменений | Уничтожаются в борьбе за существование | Отбираются, бракуются, уничтожаются |
| Направленность действия | Отбор признаков, полезных особи, популяции, виду | Отбор признаков, полезных человеку |
| Результат отбора | Новые виды | Новые сорта растений, породы животных, штаммы микроорганизмов |
| Формы отбора | Движущий, стабилизирующий, дизруптивный | Массовый, индивидуальный, бессознательный (стихийный), методический (сознательный) |
Отбор бывает массовый и индивидуальный. Массовый отбор — выделение из исходного материала целой группы особей с желательными признаками и получение от них потомства. Индивидуальный отбор — выделение отдельных особей с желательными признаками и получение от них потомства. Массовый отбор чаще применяют в селекции растений, а индивидуальный — в селекции животных, что связано с особенностями размножения растений и животных.
Систематика и происхождение[]
Особый интерес для генетиков представляет происхождение рапса. Любопытно, что в диком виде это растение не встречается. В культуре известен за 4 тыс. лет до н. э. Полагают, что рапс произошёл от скрещивания озимой или яровой сурепицы (В. campestris) с капустой огородной (В. oleracea).
Рапс (Brassica napus oleifera D.C.) относится к роду капуст (Brassica L.), семейству капустных (крестоцветных) Brassicaceae Веnс. (Cruciferae Juss.) и имеет яровую (В. napus oleifera annua Metzg.) и озимую (В. napus oleifera biennis Metzg.) формы.
Относительно места происхождения рапса до сих пор нет единого мнения. Большинство ботаников относят род Brassica и, в частности рапс, к Средиземноморскому центру происхождения культурных растений. Дикорастущий рапс неизвестен, но во многих странах Европы, Азии, Америки и Северной Африки рапс встречается в одичалом состоянии как сорняк.
По мнению Е. Н. Синской рапс происходит из Европы. Его родина -Англия и Голландия, откуда он в XVI в. Распространился в Германию, затем в Польшу и Западную Украину. В России как масличную культуру его начали возделывать с начала XIX в.
Во многих странах под названием «рапс» объединяют несколько видов этого семейства: рапс, сурепицу, горчицу сапетскую, сизию и т. д.
Рапс — естественный амфидиплоид, в происхождении которого участвовала сурепица (2n = 20, геном АА) и капуста (2n = 18 геном СC. В настоящее время насчитывают 12 разновидностей рапса. Наиболее распространенные из них — италика, россика (озимые формы) и корамбоза (яровая).
Ресинтез липидов в энтероцитах
Активация жирной кислоты
Поступившая в энтероцит (как и в любую другую клетку) жирная кислота обязательно активируется через присоединение коэнзима А. Образовавшийся ацил-SКоА участвует в реакциях синтеза эфиров холестерола, триацилглицеролов и фосфолипидов.
Реакция активации жирной кислоты
Ресинтез эфиров холестерола
Холестерол этерифицируется с использованием ацил-SКоА и фермента ацил-SКоА:холестерол-ацилтрансферазы (АХАТ).
Реэтерификация холестерола напрямую влияет на его всасывание в кровь. В настоящее время ищутся возможности подавления этой реакции для снижения концентрации ХС в крови.
Реакция ресинтеза эфиров холестерола
Ресинтез триацилглицеролов
Для ресинтеза ТАГ есть два пути:
Первый путь, основной – 2-моноацилглицеридный – происходит при участии экзогенных 2-МАГ и ЖК в гладком эндоплазматическом ретикулуме энтероцитов: мультиферментный комплекс триацилглицерол-синтазы формирует ТАГ.
Моноацилглицеридный путь образования ТАГ
Поскольку 1/4 часть ТАГ в кишечнике полностью гидролизуется, а глицерол в энтероцитах не задерживается и быстро переходит в кровь, то возникает относительный избыток жирных кислот для которых не хватает глицерола. Поэтому существует второй, глицеролфосфатный, путь в шероховатом эндоплазматическом ретикулуме. Источником глицерол-3-фосфата служит окисление глюкозы. Здесь можно выделить следующие реакции:
- Образование глицерол-3-фосфата из глюкозы.
- Превращение глицерол-3-фосфата в фосфатидную кислоту.
- Превращение фосфатидной кислоты в 1,2-ДАГ.
- Синтез ТАГ.
Глицеролфосфатный путь образования ТАГ
Ресинтез фосфолипидов
Первый путь – с использованием 1,2-ДАГ и активных форм холина и этаноламина для синтеза фосфатидилхолина или фосфатидилэтаноламина.
Ресинтез фосфолипидов из ДАГ на примере фосфатидилхолина
Второй путь – на основе синтезируемой in situ фосфатидной кислоты.
ТИПЫ СОРТОВ
Все сорта делятся на две базовые группы:
1. Местные сорта — сорта, созданные путем естественного и/или простейшего искусственного отбора в процессе возделывания культуры в конкретной местности. Такие сорта также назваются народными. Они обладают большой неоднородностью по морфологическим признакам и потребительским свойствам.









2. Селекционные сорта — сорта выведенные искусственно с помощью методов научной селекции. Они отличаются существенно большей однородностью по морфологическим признакам и свойствам.
Селекционные сорта в свою очередь по технологии селекции из исходного материала подразделяются на следующие субкатегории:
2.1. Сорт-популяция — сорт, полученный путем размножения большого числа растений, отобранных из исходного материала по заданным признакам. Эти сорта по схеме отбора эквивалентны местным, поэтому также обладают большой неоднородностью.
2.2. Линейный сорт — сорт, полученный путем размножения одного отобранного растения. Линейные сорта обладают существенно большей однородностью, чем сорта-популяции.
По технологии создания мутаций исходного материала селекционные сорта подразделяют на следующие категории:
2.A. Мутантный сорт — сорт, полученный путем отбора из популяций, полученных под воздействием мутагенных факторов. Данная технология имеет ограниченное применение в силу значительной сложности и трудоемкости.
2.B. Гибридный сорт — сорт, полученный путем гибридизации (скрещивания) двух или более сортов. Это наиболее востребованная технология селекции, поскольку она достаточно проста и эффективна, в т.ч. позволяет получать гетерозисные сорта, превосходящие по полезным свойствам родительские растения. На сегодня большинство сортов получают путем гибридизации.
2.C. Полиплоидный сорт — сорт, полученный путем кратного увеличения числа хромосом в ядре клетки, что позволяет резко улучшить качественные показатели сорта.
2.D. Генно-модифицированный сорт — сорт, полученный путем генной инженерии. В РФ сорта данного типа для культивирования не разрешены.
2.E. Сорт-клон — сорт, полученный путем размножения отобранного для сорта растения вегетативным способом — черенками, клубнями, луковицами, клубнелуковицами, корневищами и пр. При таком способе полностью сохраняется генетическая идентичность родителей и потомства, поэтому сорта — клоны в первых поколениях обладают очень высокой степенью однородности.
Внимание! Во многих популярных публикациях и даже в учебниках систематизация сортов по типам излагается недостаточно четко, в связи с чем часто допускаются ошибочные трактовки, в т.ч. противопоставление понятий сорт, селекционный сорт и сортовые культуры (семена) понятию гибрид, что неправомерно
Необходимо четко уяснить, что гибрид — это категория селекционного сорта, причем наиболее распространенная, а сорт — общее понятие, объединяющее все описанные выше типы сортов (см. определение в начале обзора). Понятие же сортовые культуры является некорректным, т.к. культуры — это культурные растения, и они обязательно характеризуются сортом, который в т.ч. подлежит обязательной регистрации в Государственном реестре селекционных достижений (см. обзор О Госсортреестре).
Сорт может быть районированным, т.е. ориентированным на определенные регионы допуска территории РФ на основании результатов проведенных сортоиспытаний. Большинство сортов, зарегистрированных в Госсортреестре, являются районированными.
Лучшие районированные сорта могут быть установлены в качестве стандартов качества для их регионов допуска. Такие сорта называются стандартными и часто указываются в характеристках близких сортов вида в качестве эталона сопоставления.
Более подробно о технологиях селекции и особенностях тех или иных категории сортов см. в специальных публикациях Библиотеки по тематике ФитоЦентра
Виды селекции растений
Различные типы существующих процессов селекции растений включают:
- инбридинг;
- обратное скрещивание;
- селекцию мутаций;
- гибридную селекцию;
- генную инженерию.
Все эти процессы включают в себя свои собственные различные методы и способы, которые способствуют повышению урожайности сельскохозяйственных культур несколькими способами.
Обратное скрещивание
При этом растение с желаемыми признаками скрещивается с растением, которое не обладает желаемыми признаками, но обладает несколькими другими признаками.
Растение, обладающее желательным признаком, например, устойчивостью к плесени, скрещивается с растением, которое не обладает этим признаком, но желательно по всем остальным признакам. Существует этап контроля качества, чтобы убедиться, что единственным изменением исходного сорта является желаемый признак.
Инбридинг
В зависимости от вида некоторые растения могут быть оплодотворены сами по себе. Это делается для получения инбредного сорта, который из поколения в поколение остается точно таким же. Поскольку он сохраняет исходные черты, он полезен тремя способами: для исследований, в качестве новых сортов с истинной селекцией и в качестве родителей гибридов.
Гибридная селекция
В этой ситуации два разных инбредных сорта скрещиваются для получения потомства со стабильными характеристиками и гибридной энергией, где потомство намного более продуктивно, чем любой из родителей.
Селекция мутаций
Мутации в генах растений приводят к появлению новых сортов.
Естественные генетические мутации существуют во всем мире. Если эти случайные примеры будут найдены и восприняты как улучшение, их можно будет использовать для создания новых сортов. В качестве альтернативы мутации можно искусственно стимулировать, подвергая растения воздействию химических веществ или радиации.
Генная инженерия
Генная инженерия помогает в создании скрещивания с желаемыми признаками, вставляя интересующую игру в ДНК культуры. Такие культуры известны как генетически модифицированные культуры. Например, Bt-культуры.
Селекция и генетика растений
Грегор Мендель (1822-84) считается «отцом генетики». Он разработал законы наследования с помощью экспериментов с гибридизацией растений. Генетика стимулировала исследования по повышению урожайности сельскохозяйственных культур за счет селекции растений.
Генетическая модификация растений достигается путем добавления выбранного гена или генов к растению или путем уничтожения гена с помощью РНК-интерференции для получения желаемого фенотипа. Растения, полученные в результате добавления гена, часто упоминаются как трансгенные растения. Если гены генетической модификации вида или скрещиваемого растения используются для проверки их нативного промотора, то они называются цисгенными растениями. Иногда генетическая модификация может привести к появлению растения с заданным признаком или признаками быстрее, чем классическая селекция, потому что основная часть генома растения не изменяется.
Современная селекция растений
Иногда множество различных генов может влиять на желаемый признак в селекции растений. Использование таких инструментов, как молекулярные маркеры или ДНК-отпечатки пальцев, может нанести на карту тысячи генов. Это позволяет селекционерам проводить скрининг больших популяций растений на предмет людей, обладающих интересующей чертой. Скрининг основывается на наличии или отсутствии определенного гена, определяемого лабораторными процедурами, а не на визуальной идентификации выраженного признака внутри растения.
Классическая или традиционная селекция растений
Традиционная селекция в значительной степени основана на гомологичной рекомбинации между хромосомами для получения генетического разнообразия. Классический селекционер растений может также использовать различные методы in vitro, такие как слияние протопластов, спасение эмбрионов или мутагенез, для получения разнообразия и получения гибридных растений, которые могут не существовать в природе.
Другой метод заключается в преднамеренном скрещивании близкородственных или отдаленных особей для получения новых сортов или линий сельскохозяйственных культур с желаемыми свойствами. Растения для скрещивания используются для введения признаков или генов одного сорта или линии в замещающий генетический фон.
Слайд 8 Снижение продуктивности и жизнеспособности организмов в результате инбридинга

первых поколениях и постепенно снижается в последующих. Этот процесс продолжается
до тех пор, пока растения не достигнут инбредного минимума, т.е. такого состояния, когда депрессия достигла наивысшего выражения и дальнейшего снижения продуктивности и жизнеспособности особей в последующих поколениях не вызывает.
3.1 Инбридинг
Инбридинг может быть как полезен, так и вреден. В процессе инбридинга депрессию вызывают аллели, понижающие жизнеспособность организмов – летальные, полулетальные, а также появление плохо приспособленных к конкретным условиям среды генотипов, которые в исходной аллогамной популяции возникаю редко, а в случае их появления элиминируют. В гетерозиготном состоянии действие рецессивных аллелей подавляется доминантными.
Слайд 7 3. ГибридизацияИнбридинг – скрещивание особей, находящихся между собой
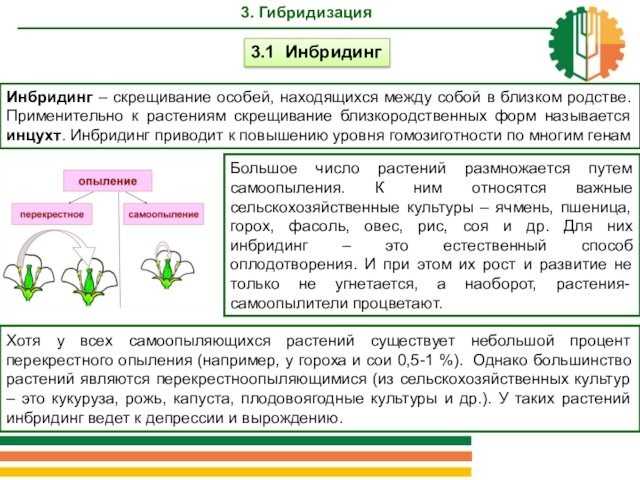
называется инцухт. Инбридинг приводит к повышению уровня гомозиготности по многим
генам
Большое число растений размножается путем самоопыления. К ним относятся важные сельскохозяйственные культуры – ячмень, пшеница, горох, фасоль, овес, рис, соя и др. Для них инбридинг – это естественный способ оплодотворения. И при этом их рост и развитие не только не угнетается, а наоборот, растения-самоопылители процветают.
3.1 Инбридинг
Хотя у всех самоопыляющихся растений существует небольшой процент перекрестного опыления (например, у гороха и сои 0,5-1 %). Однако большинство растений являются перекрестноопыляющимися (из сельскохозяйственных культур – это кукуруза, рожь, капуста, плодовоягодные культуры и др.). У таких растений инбридинг ведет к депрессии и вырождению.
Слайд 18 Методы преодоления нескрещиваемости отдаленных гибридов 1. Применение реципрокных

пестика различных видов, а также разным уровнем плоидности эндосперма Так,
при получении гибридов редьки и капусты материнским растением является редька. Обратный гибрид получить никому не удалось. При получении тритикале в вариантах, где материнским растением была пшеница, а опылителем рожь, завязываемость гибридных семян составляла 60,5 %, в обратном скрещивании – 3,6 %.








2. Изменение уровня плоидности родителей. Перевод скрещиваемых видов на тетраплоидный уровень способствует получению гибридных семян
4. Причины нескрещиваемости и бесплодия отдаленных гибридов и методы их преодоления
Слайд 47 Гаплоид – организм, имеющий в соматических клетках гаметический

покрытосеменных растений, относящихся к 75 родам и 33 семействам. Они
характеризуются уменьшенным размером клеток и органов
В фенотипе гаплоидов проявляются рецессивные гены, так как их не перекрывают доминантные аллели. Гаплоиды отличаются пониженной жизнеспособностью из-за влияния рецессивных генов, особенно у перекрестноопыляющихся культур
В фенотипе гаплоидов проявляются рецессивные гены, так как их не перекрывают доминантные аллели. Гаплоиды отличаются пониженной жизнеспособностью из-за влияния рецессивных генов, особенно у перекрестноопыляющихся культур
7. Полиплоидия
Слайд 31 6. Соматическая гибридизацияСоматическую гибридизацию применяют для получения отдаленных

гибридные клетки были получены при слиянии клеток разных линий мышей
(внутривидовые гибриды). Межвидовые гибриды получены между клетками человека и мыши, мыши и хомячка, мыши и цыпленка
Селекция гибридных клеток и регенерация у животных
Клетки животных помещают на искусственные питательные среды, добавляют специфические химические вещества (например, полиэтиленгликоль) или инактивированные вирусы (например, вирус Сендай)
При этом происходит слияние плазматических мембран клеток и образуется клетка с двумя ядрами – дикарион
Далее ядра сливаются, образуется клетка с хромосомами обоих родителей – синкарион
Синкарионы переносят для пролиферации на селективные питательные среды. При первых делениях клетки происходит потеря хромосом одного из видов. Клеточная линия, хромосомы которой утрачиваются после слияния, называется донорной, другая – реципиентной
Слайд 51 Контрольные вопросы3. Какие методы применяют для преодоления нескрещиваемости

какие методы применяют для преодоления стерильности гибридов первого поколения? 5.
Что называют синтезом и ресинтезом видов? Приведите примеры.
6. Назовите этапы соматической гибридизации и приведите примеры применения соматической гибридизации у растений и животных организмов
1. Дайте определение понятию «отдаленная гибридизация»
2. Каковы причины нескрещиваемости отдаленных видов?
9. Что называют полиплоидным рядом? Приведите примеры полиплоидных рядов.
10. Какие организмы называют автополиплоидными, аллоплоидными, анеуплоидными, гаплоидными?
7. Что называют полиплоидией? Приведите классификацию полиплоидов.
8. Дайте определение понятию основное число хромосом.