
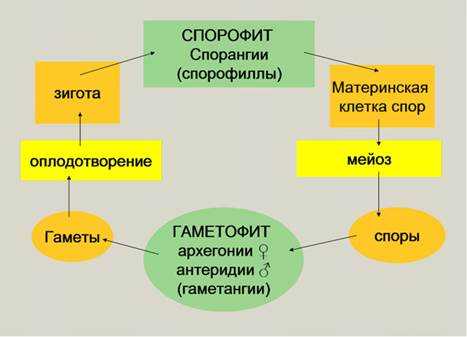
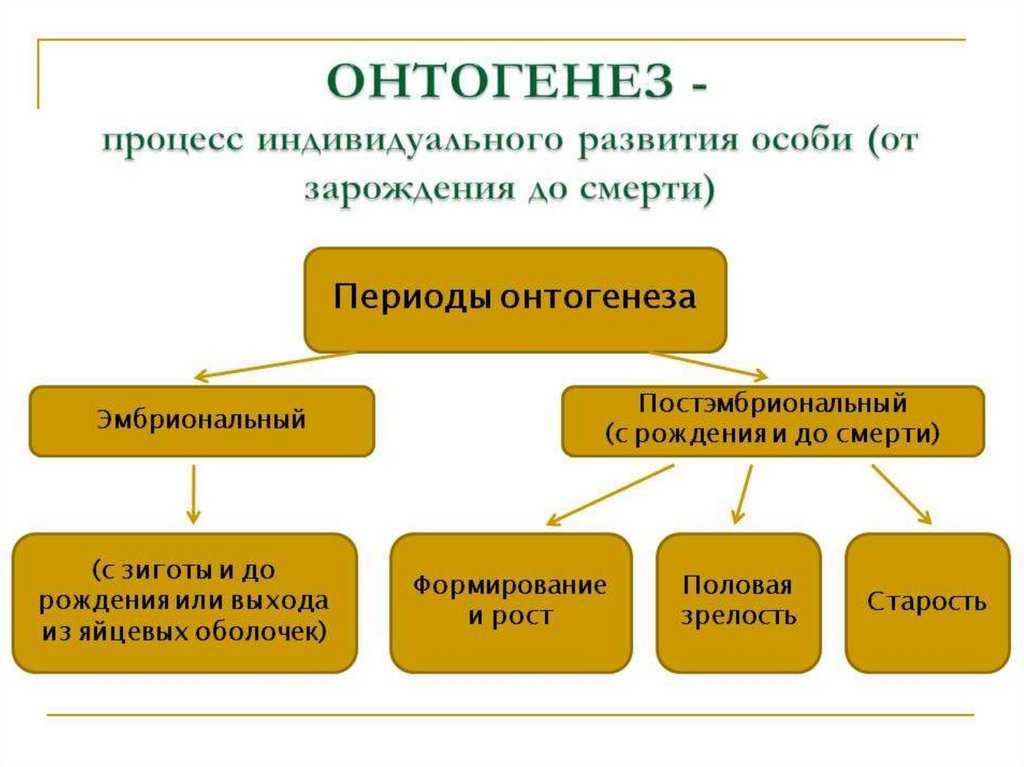
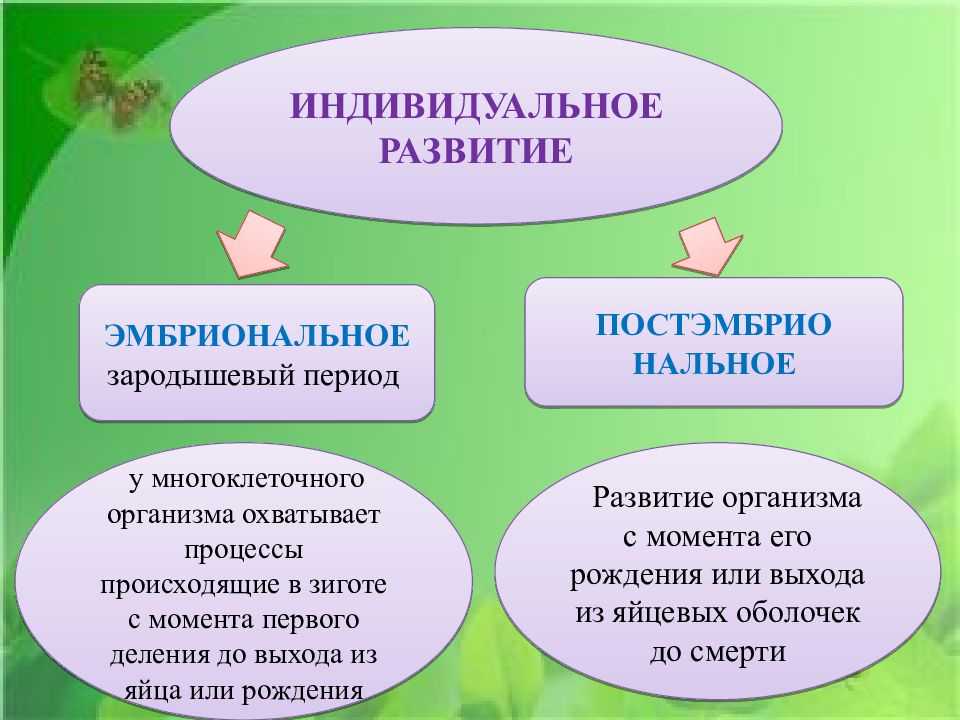
Спорогенез и гаметогенез у растенийСтраница 1
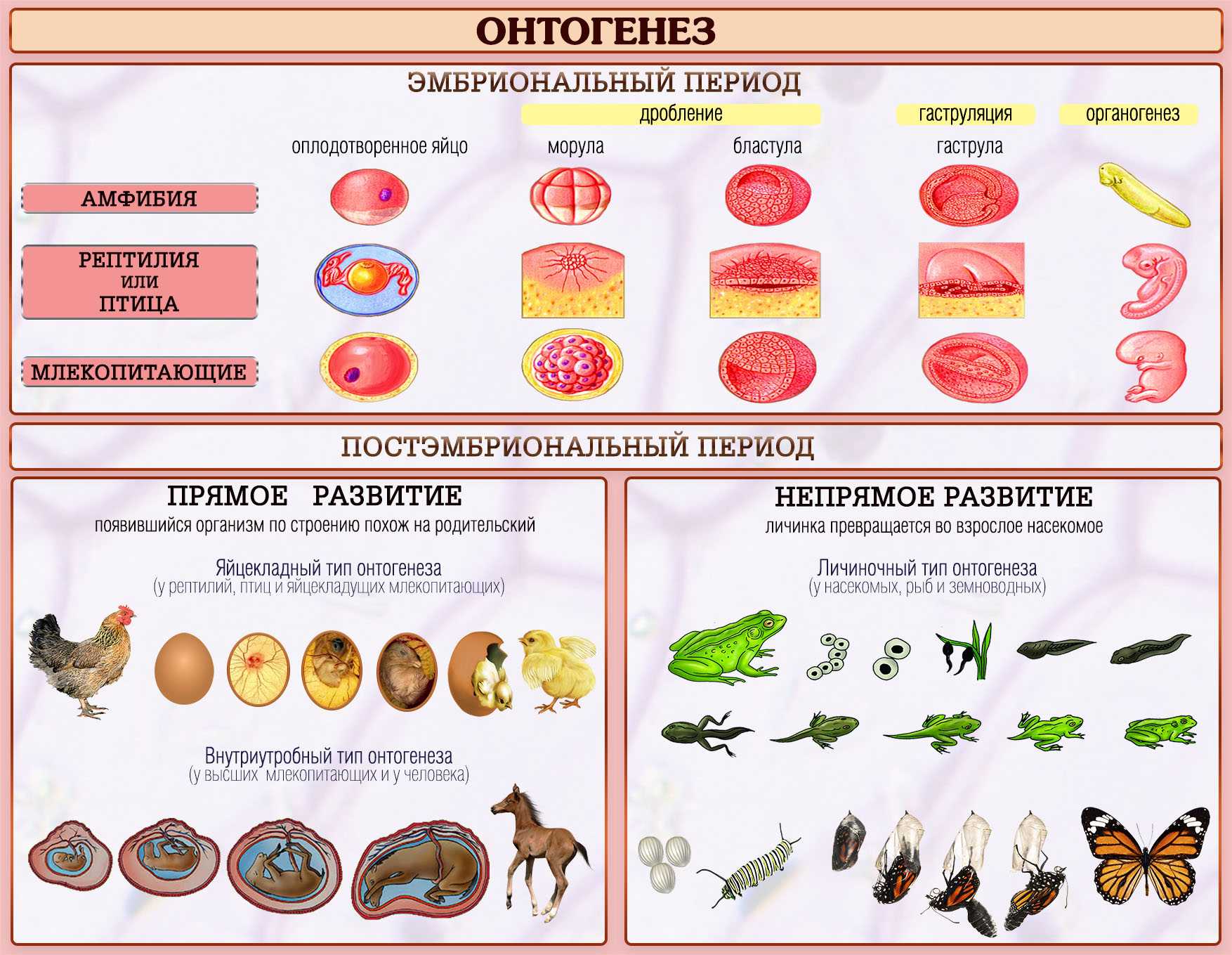
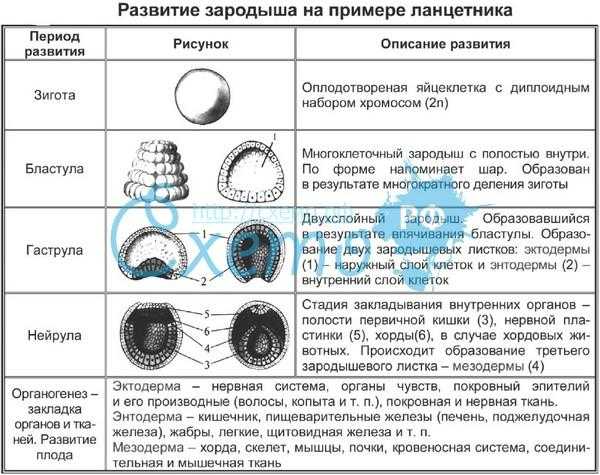
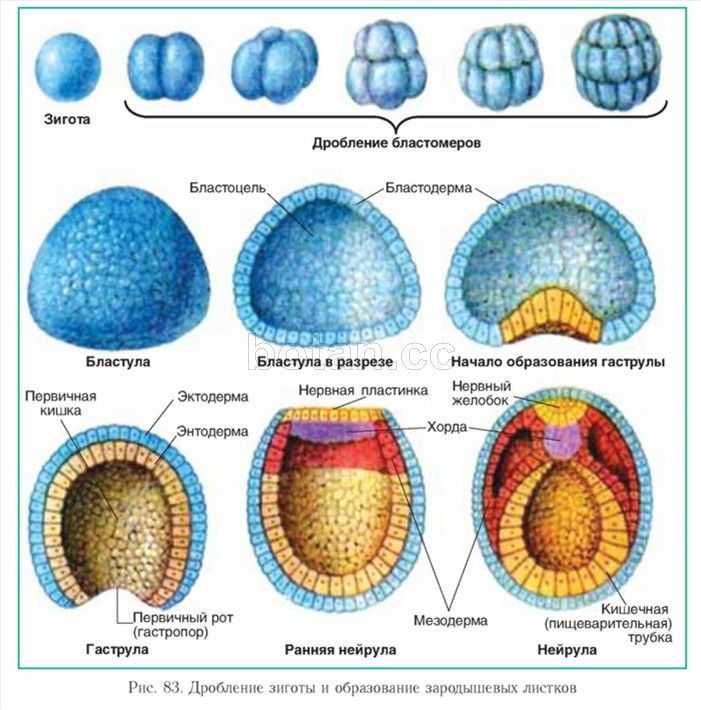
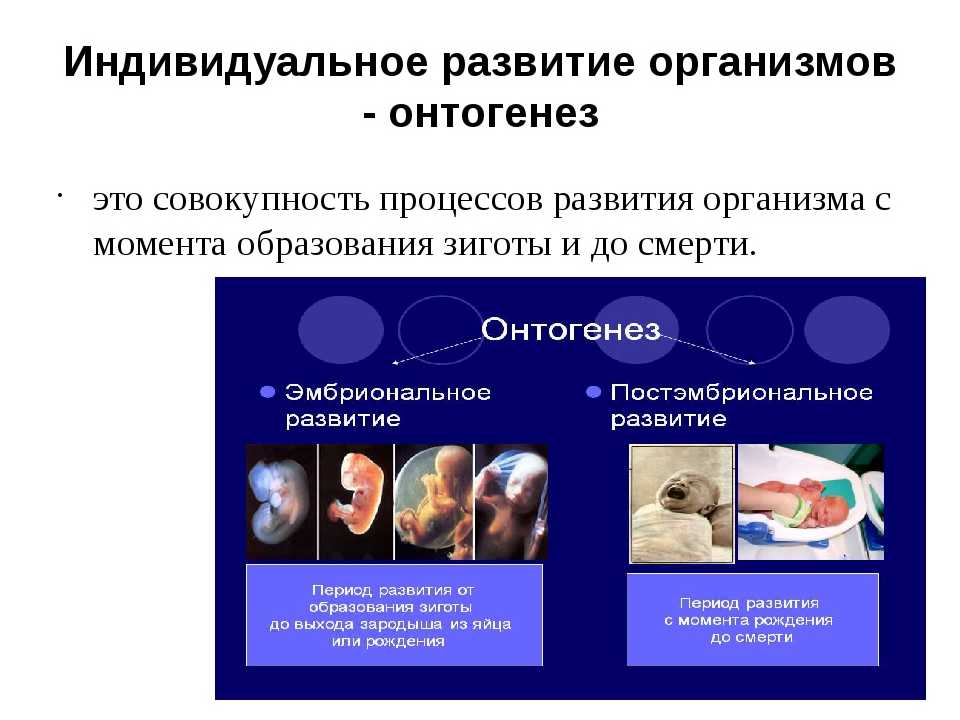
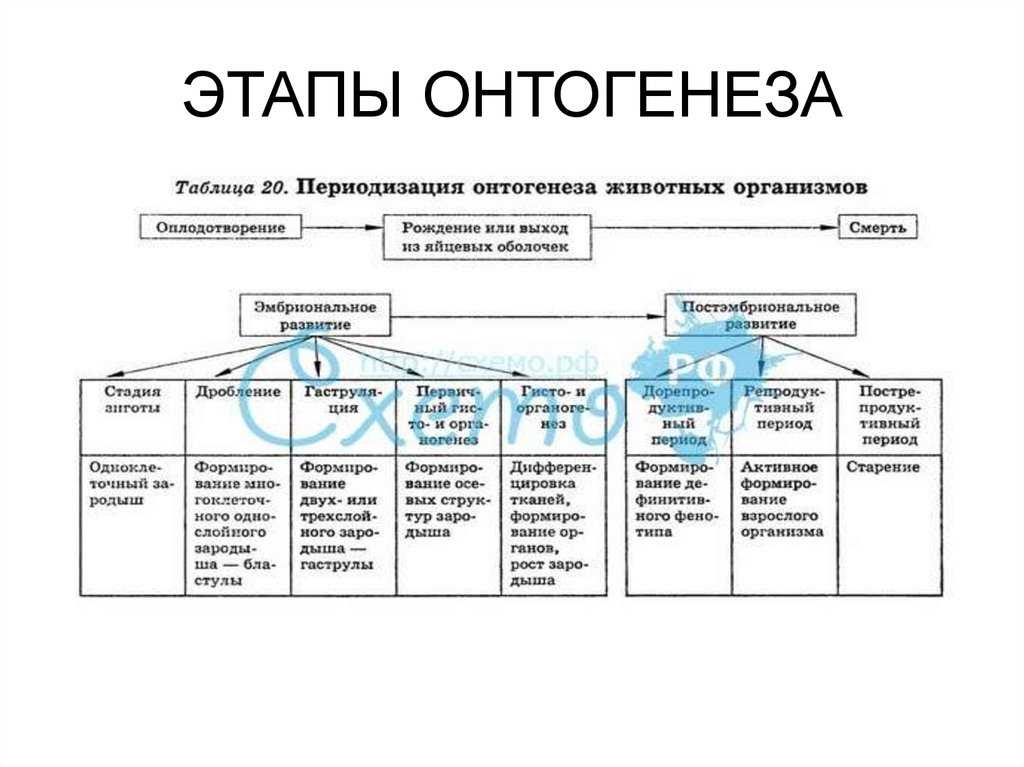
Способность к размножению, или самовоспроизведению, является одним из обязательных и важнейших свойств живых организмов. Размножение поддерживает длительное существование вида, обеспечивает преемственность между родителями и их потомством в ряду многих поколений. Оно приводит к увеличению численности особей вида и способствует его расселению. У растений, подавляющее большинство которых ведет прикрепленный образ жизни, расселение в процессе размножения — единственный способ занять большую территорию обитания.
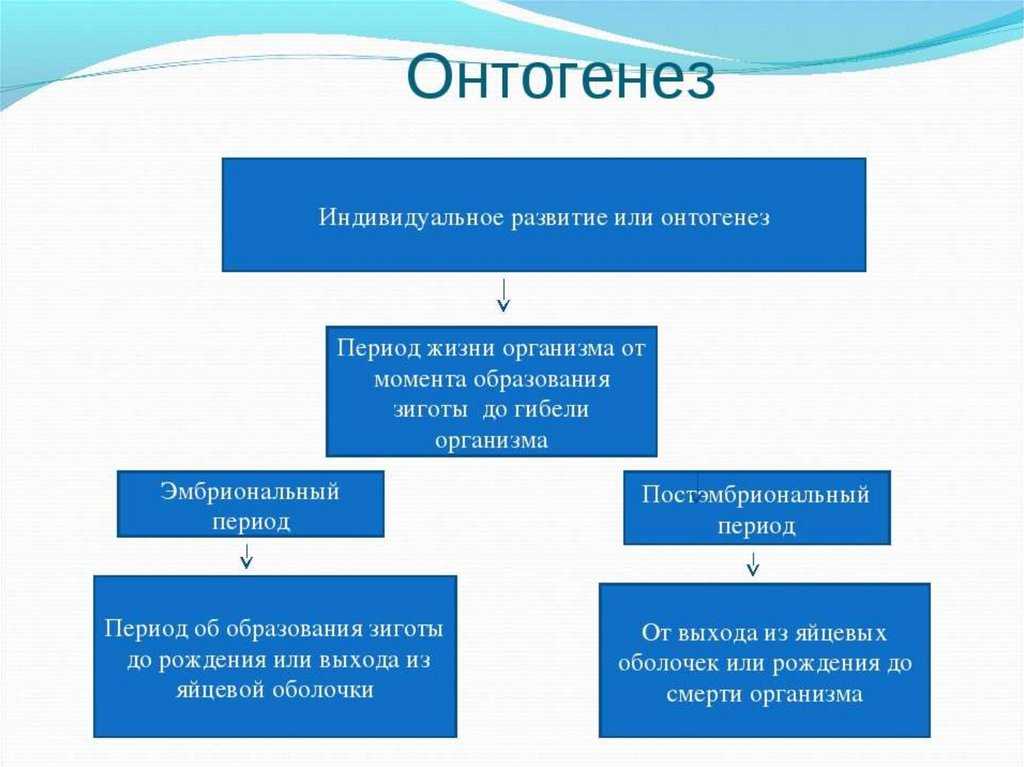
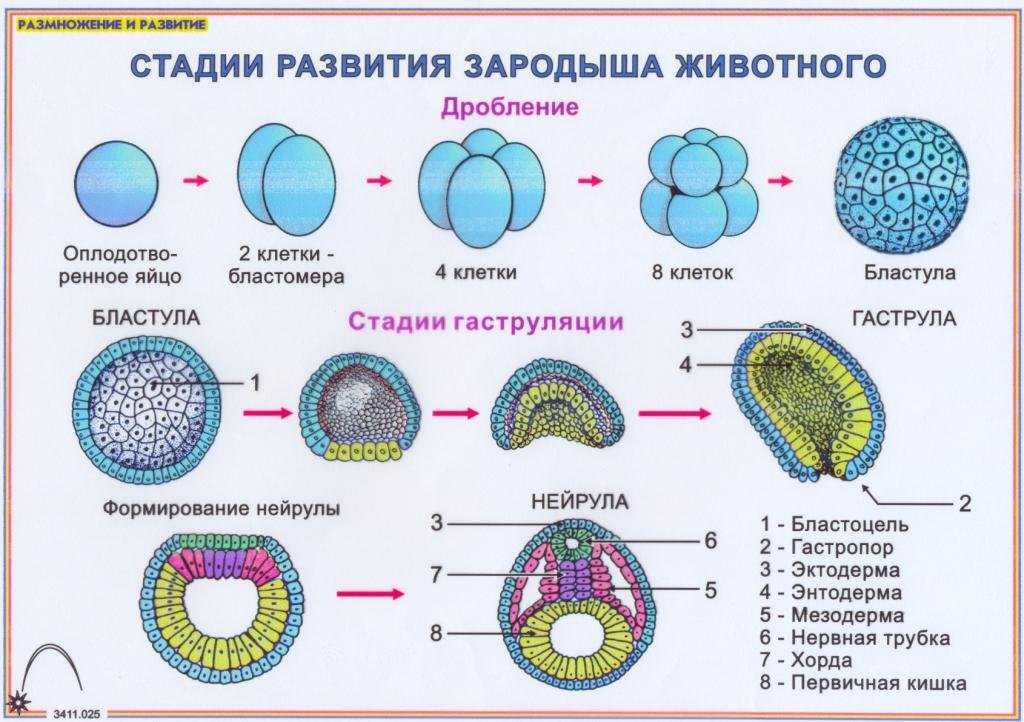
Различают два типа размножения: бесполое и половое. В, бесполом размножении участвует только одна родительская особь, которая делится, почкуется и образует споры. Размножение при помощи вегетативных органов у растений называется вегетативным. В случае полового размножения особи нового поколения появляются при участии двух организмов — материнского и отцовского.
Вегетативное размножение основано на способности организмов восстанавливать (регенерировать) недостающие части. Этот способ размножения широко распространен в природе, но с наибольшим разнообразием оно осуществляется у растений, особенно у цветковых.
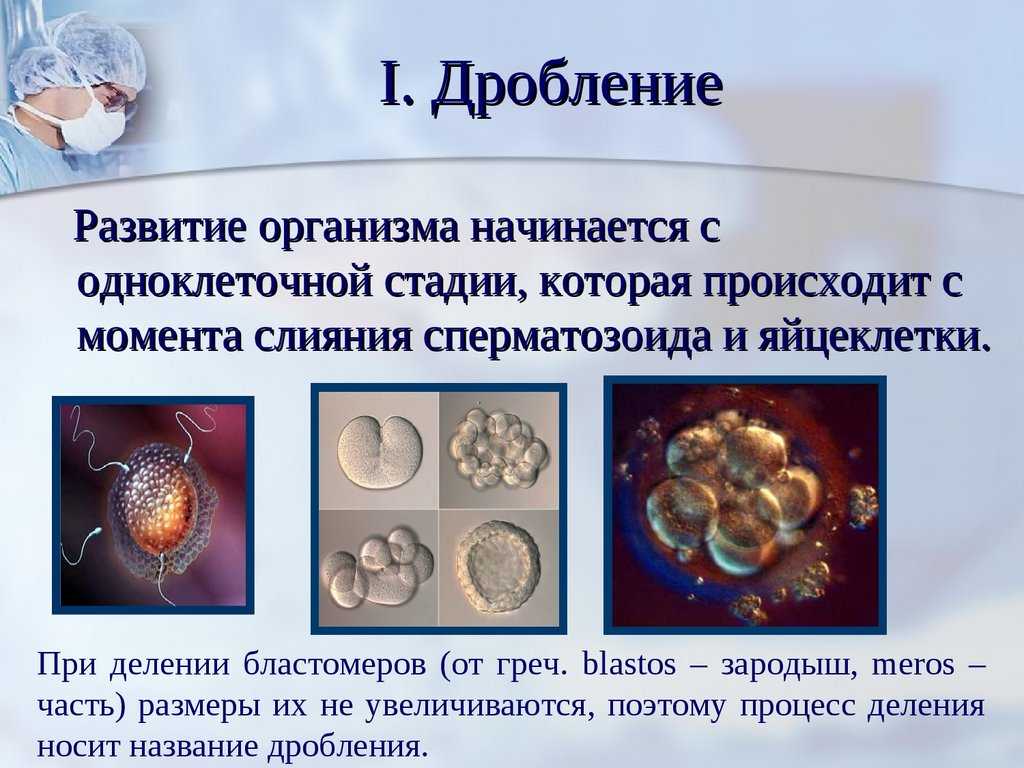
Бесполое размножение характеризуется тем, что для воспроизводства потомства образуются специализированные клетки — споры, каждая из которых прорастает и дает начало новому организму. Спорообразование встречается у простейших (малярийный плазмодий), грибов, водорослей, мхов, плаунов, хвощей и папоротников. У голо — и покрытосеменных растений споры непосредственно в процессе, размножения не участвуют.
Споры образуются путем митоза или мейоза в обычных вегетативных клетках материнского организма или специальных органах — спорангиях и представляют собой микроскопические одноклеточные образования.
При любой форме бесполого размножения — частями тела или спорками — наблюдается увеличение численности особей данного вида без повышения их генетического разнообразия: все особи являются точной копией материнского организма. Эта особенность используется человеком для получения однородного, с хорошими признаками, потомства у плодово-ягодных, декоративных и других групп растений. Новые признаки у таких организмов появляются только в результате мутаций.
Половое размножение существенно отличается от бесполого тем, что в данном случае генотип потомков возникает в результате перекомбинации генов, принадлежащих обоим родителям. Это повышает возможности организмов в приспособлении к меняющимся условиям среды.
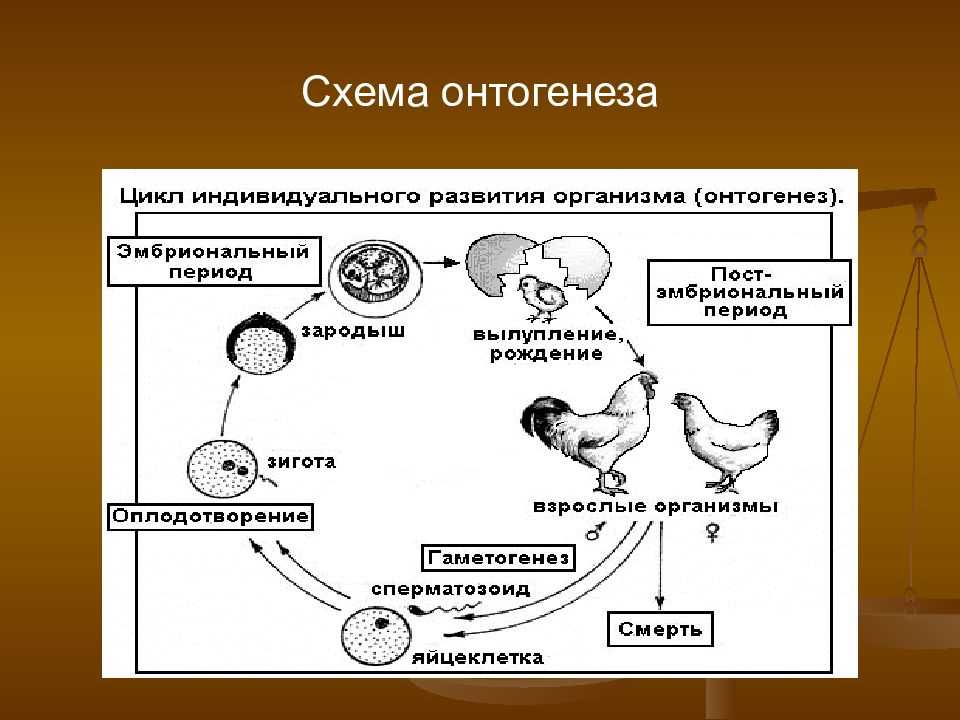
Половое размножение характеризуется наличием полового процесса, одним из важнейших этапов которого является слияние половых клеток, или гамет, специализированных гаплоидных клеток, одетых плазматической мембраной. Гаметы различаются по строению и физиологическим свойствам и делятся на мужские (подвижные — сперматозоиды, неподвижные — спермин) и женские (яйцеклетки). В отличие от спор одна гамета, за исключением случаев партеногенеза, не может дать начало новой особи. Этому предшествует процесс слияния двух половых клеток — оплодотворение, в результате которого образуется зигота. В дальнейшем из зиготы развивается зародыш нового организма.
Страницы: 1
Анализ
МОЛЕКУЛЯРНЫЙ АНАЛИЗ, установление качественного и количественного состава хим. соединений и их смесей.
При качественном анализе смеси хим. соединений обычно предварительно разделяют различными методами (хроматографией, ректификацией, кристаллизацией, экстракцией, осаждением, термической диффузией и др.); затем для разделенных в-в опред …
Неолит
Первая значительная перемена во взаимоотношениях человека с природным окружением происходит тогда, когда человек начинает заниматься скотоводством и земледелием. С этого момента прекращается исключительное влияние природы на человеческое общество; оно становится взаимным.
Прирост населения начинается в неолите при переходе к производящ …
Характеристика семейств: розоцветные и яснотковые
Розоцветные
Rosaceae
Яснотковые
Lamiaceae
Состав
Жирное масло, цианогенные гликозиды, Тритерпеновые сапонины, танниды, терпеноиды, полисахариды, редко алколоиды, кумарины.
Эфирные масла, ди — и тритерпеноиды, сапонины, полифенолы и танниды, иридоиды, хиноны, кумарины, гормоны линьки насекомых, редко алколоиды.
…
Бесплодие отдаленных гибридов, его причины и способы преодоления
Так как одним из методов селекции является гибридизация, то большую роль играет выбор типа скрещиваний, т.е. система скрещиваний.
Системы скрещивания могут быть разделены на два основных типа: близкородственное (инбридинг — разведение в себе) и скрещивание между неродственными формами (аутбридинг — неродственное разведение). Если принудительное самоопыление приводит к гомозиготизации, то неродственные скрещивания — к гетерозиготизации потомков от этих скрещиваний.
Инбридинг, то есть принудительное самоопыление перекрестноопыляющихся форм, кроме прогрессирующей с каждым поколением степени гомозиготности, приводит и к распадению, разложению исходной формы на ряд чистых линий. Такие чистые линии будут обладать пониженной жизнеспособностью, что, по-видимому, связано с переходом из генетического груза в гомозиготное состояние всех рецессивных мутаций, которые в. основном являются вредными.
Чистые линии, полученные в результате инбридинга, имеют различные свойства. У них различные признаки проявляются по-разному. Кроме того, различна и степень снижения жизнеспособности. Если эти чистые линии скрещивать между собой, то, как правило, наблюдается эффект гетерозиса.
Гетерозис — явление повышенной жизнеспособности, урожайности, плодовитости гибридов первого поколения, превышающих по этим параметрам обоих родителей. Уже со второго поколения гетерозисный эффект угасает. Генетические основы гетерозиса не имеют однозначного толкования, но предполагается, что гетерозис связан с высоким уровнем гетерозиготности у гибридов чистых линий (межлинейные гибриды). Производство чистолинейного материала кукурузы с использованием так называемой цитоплазм этической мужской стерильности было широко изучено и поставлено на промышленную основу в США. Ее использование исключало необходимость кастрировать цветки, удалять пыльники, так как мужские цветки растений, используемые в качестве женских, были стерильны.
Разные чистые линии обладают разной комбинационной способностью, то есть дают неодинаковый уровень гетерозиса при скрещиваниях друг с другом. Поэтому, создав большое количество чистых линий, экспериментально определяют наилучшие комбинации скрещиваний, которые затем используются в производстве.
Отдаленная гибридизация — это скрещивание растений, относящихся к различным видам. Отдаленные гибриды, как правило, стерильны, что связано с содержанием в геноме различных хромосом, которые в мейозе не конъюгируют. В результате этого формируются стерильные гаметы. Для устранения данной причины в 1924 г. советским ученым Г.Д. Карпеченко было предложено использовать удвоение числа хромосом у отдаленных гибридов, которое приводит к образованию амфидиплоидов по следующей схеме:
Р: пшеница Х рожь
Р: пшеница Х рожь
2n = 42 2n = 14
(21 пара) (7 пар)
G:
2. Норма реакции генотипа
При формировании генетических представлений о связи между геном и признаком изначально предполагалось, что каждому признаку соответствовал особый детерминант (наследственный фактор), который обусловливал развитие своего признака. Однако такие представления далеки от истины, а прямые и однозначные связи гена с признаком на самом деле скорее исключение, чем правило. Было установлено, что на один признак могут влиять многие гены и, наоборот, один ген часто влияет на многие признаки. Кроме того, действие гена может быть изменено соседством других генов или условиями внешней среды.
В онтогенезе действуют скорее не отдельные гены, а весь генотип как целостная интегрированная система со сложными связями и взаимодействиями ее компонентов. Более того, эта система не является застывшей, она динамична, меняется, совершенствуется во времени, в результате генных мутаций постоянно появляются новые гены. Могут формироваться также качественно новые хромосомы за счет хромосомных мутаций и даже новые геномы за счет геномных мутаций. Вновь возникшие гены могут сразу же вступать во взаимодействие с уже имевшимися генами или менять, модифицировать характер работы последних, даже будучи рецессивными, т.е. не проявляясь сами по себе.




Таким образом, в каждый конкретный промежуток времени у каждого вида растений и животных генотип проявляет себя как исторически сложившаяся к данному моменту целостная система.
Характер проявления действия гена может изменяться в различных ситуациях и под влиянием различных факторов. Законы Менделя отражают законы наследования, то есть передачи генов в ряду поколений, только при обязательном соблюдении двух условий: гены должны быть локализованы в разных парах гомологичных хромосом (это дает им возможность независимо комбинироваться и наследоваться) и за каждый признак должен отвечать только один ген. Однако это далеко не всегда так. Для того, чтобы убедиться в том, что характер проявления генов разнообразен, рассмотрим свойства генов и особенности их проявления в признаках:
ген дискретен в своем действии, то есть, прерывист, обособлен в своей активности от других генов;
ген специфичен в своем проявлении, т.е. отвечает за строго определенный признак (на молекулярном уровне каждый ген отвечает за синтез одного конкретного белка);
ген может действовать градуально, то есть может усиливать степень проявления признака (например, увеличивать количество синтезируемого вещества) при увеличении числа доминантных аллелей (дозы гена);
один ген может влиять на развитие разных признаков — это множественное, или плейотропное, действие гена;
разные гены могут оказывать одинаковое действие на развитие одного и того же признака — это множественные гены, или полигены; при этом чаще всего наблюдается усиление или ослабление признаков — в таком случае это кумулятивное (накопительное) действие гена, которое обусловливает проявление так называемых количественных признаков;
ген может вступать во взаимодействие с другими генами, что приводит к появлению новых признаков. Поскольку гены дискретны и специфичны, они взаимодействуют не непосредственно, а продуктами своих реакций — веществами, синтезированными под их контролем;
действие гена может быть модифицировано изменением его местоположения в хромосоме (эффект положения) или условиями внешней среды и другими факторами.
Множественное действие генов — это способность гена воздействовать на несколько признаков одновременно.
В процессе индивидуального развития организма фенотип может меняться, а генотип остается таким же, каким был получен от родителей при слиянии их гамет (процесс мутирования в данном случае во внимание не принимается). Как правило, роль генотипа в определении фенотипа является решающей
Это относится в первую очередь к проявлению ряда качественных признаков (красная окраска цветков, желтая и зеленая окраска семян гороха, голубой цвет глаз у человека, наличие ушной раковины и т.д.), а также к большинству простых биохимических признаков (синтез определенных специфических белков при наличии всех необходимых компонентов).
Список литературы
1.
Алтухов Ю.П. Генетические процессы в популяциях. — М.: Мир, 1983.
2.
Биология. / Сост. Лемеза Н.А., Морозик М.С., Морозов Е.И. — Минск: Университетское,
1999.
3.
Дубинин Н.П. Общая генетика. — М.: Колос, 1987.
4.
Замотайлов С.С., Бурдун А.М. Краткий курс генетики. — М.: Агропромиздат,
1987.
5.
Основы генетики и селекции. / Под ред. Дубникова А.С. — М.: Колос, 1997.
Основы генетики и селекции. /
Под ред. Дубникова А.С. — М.: Колос, 1997.
Алтухов Ю. П.Генетические
процессы в популяциях. — М.: Мир, 1983.
Замотайлов С. С., Бурдун А.
М.Краткий курс генетики. — М.: Агропромиздат, 1987.
Дубинин Н. П.Общая
генетика. — М.: Колос, 1987.
Биология. / Сост. Лемеза Н.А.,
Морозик М.С., Морозов Е.И. – Минск: Университетское, 1999. – с.-142.
Микроспорогенез и микрогаметогенез
Мы рассмотрим микроспорогенез и микрогаметогенез на примере покрытосеменных растений как наиболее общем. В субэпидермальной ткани молодого пыльника обособляется специальная спорогенная ткань, называемая археспорием. Каждая первичная археспориальная клетка после ряда делении становится материнской клеткой пыльцы (микроспороцитом), которая проходит все фазы мейоза.
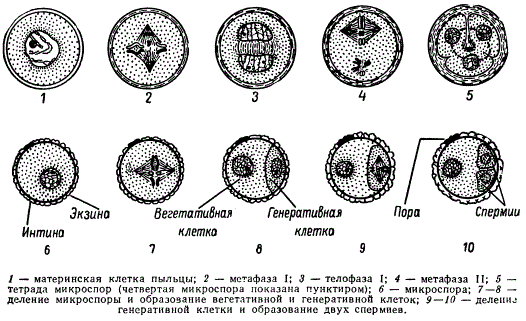
Схема микроспорогенеза (1-6) и микрогаметогенеза (5-10) у растений
В результате двух мейотических делений возникают четыре гаплоидные микроспоры. Последние лежат четвёрками и называются клеточными тетрадами.

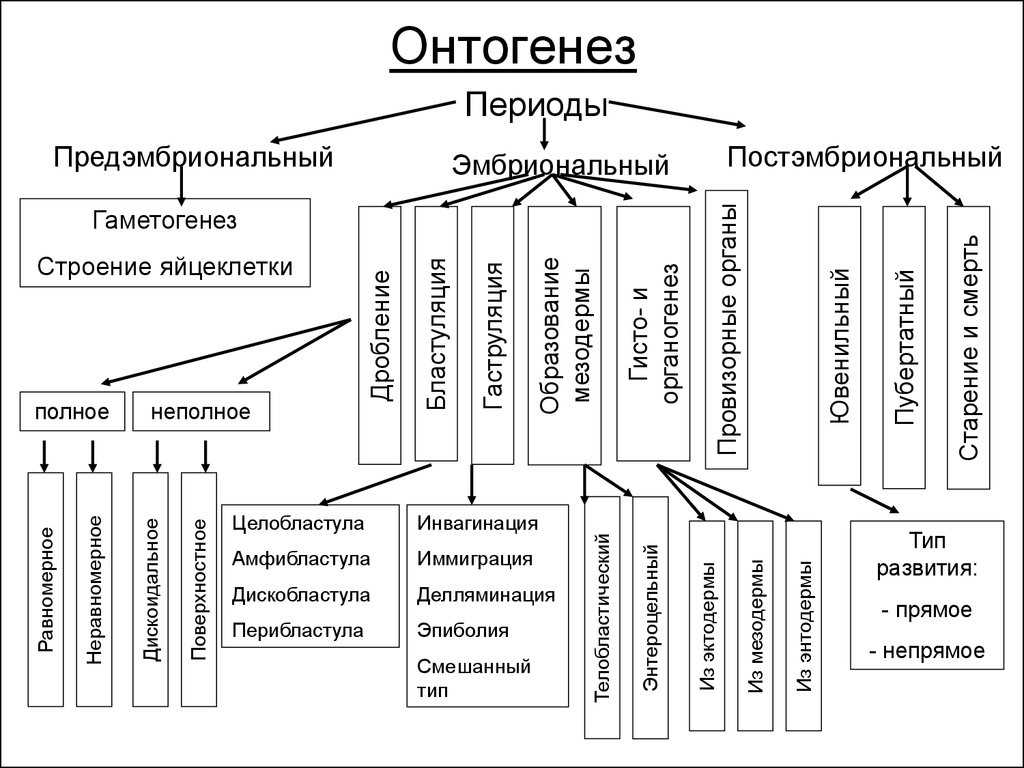


У однодольных растений каждое деление ядра в мейозе, как правило, сопровождается цитокинезом; у двудольных оба деления клетки наступают одновременно по окончании мейоза.
При созревании клеточные тетрады распадаются на отдельные микроспоры с образованием внутренней (интина) и наружной (экзина) оболочек. Наружная оболочка, как правило, грубая, кутинизированная, поверхность ее либо гладкая, либо шероховатая; приспособленная для переноса пыльцы и прилипания ее к рыльцу пестика. Этим заканчивается микроспорогенез вслед за образованием одноядерной микроспоры начинается микрогаметогенез. Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и ее ядро не делятся. В ней накапливаются запасные питательные вещества, который в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться еще в пыльцевом зерне или в процессе его прорастания в пыльцевой трубке. В результате образуются две мужские половые клетки, которые в отличие от сперматозоидов животных называются спермиоклетками, или спермиями.
Таким образом, из одной споры с гаплоидным набором хромосом в результате двух митотических делений образуются три ядра: Два из них — спермии и одно — вегетативное. При образовании пыльцевой трубки это вегетативное ядро в полужидком диффузном состоянии переходит в пыльцевую трубку.
Процесс деления генеративной клетки и образование спермиев в пыльцевой трубке были впервые подробно изучены С. Г. Навашиным в 1910 г. на лилейных растениях.
Справочная информация
ДокументыЗаконыИзвещенияУтверждения документовДоговораЗапросы предложенийТехнические заданияПланы развитияДокументоведениеАналитикаМероприятияКонкурсыИтогиАдминистрации городовПриказыКонтрактыВыполнение работПротоколы рассмотрения заявокАукционыПроектыПротоколыБюджетные организацииМуниципалитетыРайоныОбразованияПрограммыОтчетыпо упоминаниямДокументная базаЦенные бумагиПоложенияФинансовые документыПостановленияРубрикатор по темамФинансыгорода Российской Федерациирегионыпо точным датамРегламентыТерминыНаучная терминологияФинансоваяЭкономическаяВремяДаты2015 год2016 годДокументы в финансовой сферев инвестиционной
8.
Какой хромосомный набор характерен для листьев растения
кукушкина льна, его гамет и спорогона (коробочка на ножке)?
Объясните результаты в каждом случае.
Для листьев мха кукушкина льна характерна плоидность n,
так как взрослое растение – гаметофит, на котором
развиваются листья, образуется при прорастании споры
плоидностью n за счёт митозов.
Гаметы мха имеют плоидность n, так как они образуются в
половых органах мха – архегониях и антеридиях, которые
находятся на гаплоидном гаметофите. Гаметы образуются
митозами.
Спорогон на ножке – это спорофит, бесполое поколение,
которое образуется за счет митозов зиготы. Зигота имеет
плоидность 2n, следовательно, клетки спорогона имеют
плоидность 2n.
Бизнес и финансы
БанкиБогатство и благосостояниеКоррупция(Преступность)МаркетингМенеджментИнвестицииЦенные бумагиУправлениеОткрытые акционерные обществаПроектыДокументыЦенные бумаги — контрольЦенные бумаги — оценкиОблигацииДолгиВалютаНедвижимость(Аренда)ПрофессииРаботаТорговляУслугиФинансыСтрахованиеБюджетФинансовые услугиКредитыКомпанииГосударственные предприятияЭкономикаМакроэкономикаМикроэкономикаНалогиАудитМеталлургияНефтьСельское хозяйствоЭнергетикаАрхитектураИнтерьерПолы и перекрытияПроцесс строительстваСтроительные материалыТеплоизоляцияЭкстерьерОрганизация и управление производством
Развитие половых клеток у цветковых растений
У
растений наблюдается регулярная смена ядерных фаз (гаплоидной и диплоидной).
Особого внимания заслуживают цветковые растения – самые распространенные на
Земле. В жизненном цикле высших растений выделяют смену двух поколений:
гаметофита и спорофита.Гаметофит– небольшое растеньице полового
поколения, на котором формируются половые органы, продуцирующие гаметы. На нем
развиваются как женские, так и мужские гаметы. У семенных растений гаметофиты
практически утратили способность к самостоятельному существованию.
Преобладающим поколением являетсяспорофит(большинство клеток диплоидны), обычно
представляющий собой крупное листостебельное растение, существующее достаточно
длительный срок. Спорофит образуется после слияния мужских и женских гаплоидных
гамет.
Цветок
– основной орган размножения покрытосеменных цветковых растений. Цветок можно
считать как спорофитом, органом бесполого размножения (так как он производит
микроспоры и мегаспоры), так и гаметофитом – органом полового размножения (так
как из микроспор развиваются мужские гаметы– спермии, а из мегаспор – женские –
яйцеклетки).
Развитие
пыльцевых зерен происходит в пыльцевых гнездах –микроспорангияхпыльников – в два этапа.
Этап
первый –микроспорогенезспорогенной
ткани делятся митозом, образуя клетки микроспор –микроспороциты(2n). Микроспороциты делятся
мейозом, образуя микроспоры(n).Каждая материнская клетка дает четыре микроспоры (тетрада микроспор).
Этап
второй –микрогаметогенез– развитие микрогаметофита. Каждая микроспора
(n) делится митозом, образуямикрогаметофит– мужской гаметофит, илипыльцевое зерно.Сначала осуществляется процесс
бесполого размножения спорофита, для чего и используются мелкие споры. Затем
внутри пыльцевого мешка из прорастающей (делящейся) споры формируется
микроскопический мужской гаметофит, являющийся уже новым половым поколением.
Развитиезародышевого мешка происходит в семязачатке (мегаспорангии) в два
этапа. Первый этап –мегаспорогенез– развитие мегаспор. Спорогенные
клетки (2n) делятся митозом, образуя клетки мегаспор –мегаспороциты(2n). Мегаспороциты делятся
мейозом, образуямегаспоры(n). Каждая материнская клетка
дает четыре мегаспоры. В мегагаметофите развивается только одна из микроспор(обычно нижняя), остальные дегенерируют. Второй этап –метагаметогенез– развитие мегагаметофита
(зародышевого мешка). Оставшаяся из четырех однамегаспора(n) последовательно делится тремя
митозами без цитокинеза (делятся только ядра). Образуется по четыре ядра на
полюсах зародышевого мешка –восьмиядерный
зародышевый мешок.
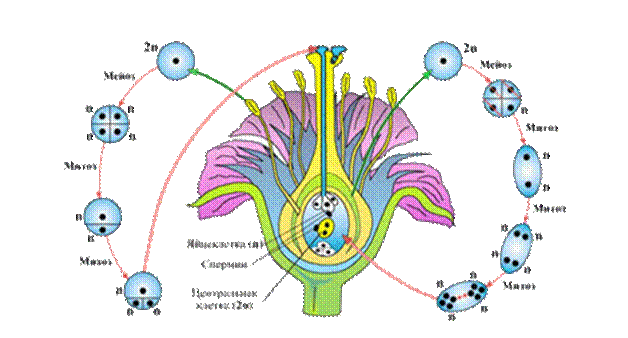
Два
ядра от полюсов отходят к центру исливаются вместе, образуя центральные (вторичные) ядра (2n). Остающиеся
на полюсах ядра превращаются в клетки:антиподы(n), яйцеклетку(n),синергиды(n). Формируется мегагаметофит
(зародышевый мешок).
Необходимо
обратить внимание на тот факт, что у высших растений (в отличие от животных)
процесс образования половых клеток осуществляется с помощью митоза. У всех
многоклеточных животных и человека для этого используется мейоз
Мужской гаметофит
у цветковыхрастений
состоит из 3 клеток, при этом один спермий оплодотворяет яйцеклетку
зародышевого мешка, а другой – центральную яйцеклетку. Происходит «двойное
оплодотворение» (его открыл русский цитолог и эмбриолог растенийС.Г. Навашин
первый спермий (n) + яйцеклетка (n) = зигота (2n);
второй спермий (n) + центральное ядро (2n) = первичное ядро
эндосперма (Зn).
Результатом
полового размножения гаметофита цветкового растения является образование
диплоидной зиготы и крупной триплоидной клетки. Их деление путем митоза в
конечном итоге приводит к формированию зародыша и эндосперма семени (запасы
питательных веществ). Семя – это важный этап в развитии нового поколения
спорофита.
Спорогенез, гаметогенез и оплодотворение у растений
18 ноября, 2009
У Покрытосеменных растений спорогенез, гаметогенез и оплодотворение представляют собой непрерывную последовательность репродуктивных процессов, происходящих в цветках. В типичных цветках репродуктивные органы представлены гинецеем (совокупностью плодолистиков, образующих женскую репродуктивную сферу) и андроцеем (совокупностью тычинок, образующих мужскую репродуктивную сферу).
Мегаспорогенез, образование зародышевого мешка и яйцеклетки
Мегаспорогенез протекает в женской репродуктивной сфере – в гинецее. Морфологически гинецей представлен пестиком (или несколькими пестиками). В состав отдельного пестика входят: завязь, столбик и рыльце. Внутри завязи находятся семязачатки (один или несколько). Внутреннее содержимое семязачатка называется нуцеллус; снаружи семязачаток окружен покровами (одиночным или двойным интегументом). В нуцеллусе семязачатка (2n) содержится одна археспориальная клетка, способная делиться путем мейоза (у некоторых растений археспорий многоклеточный). В результате мейоза из археспориальной клетки (материнской клетки мегаспор) образуется четыре мегаспоры. Вскоре три из них отмирают, а одна увеличивается в размерах и трижды делится путем митозов. В результате образуется восьмиядерный зародышевый мешок (женский гаметофит). Три ядра вместе с прилегающей цитоплазмой образуют клетки-антиподы, два ядра – одно центральное диплоидное ядро; два ядра – две клетки-синергиды; одно ядро становится ядром яйцеклетки.

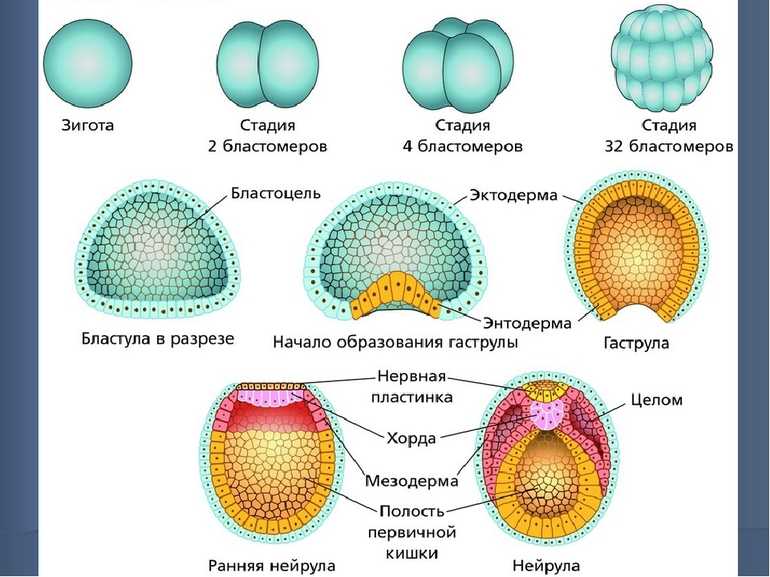
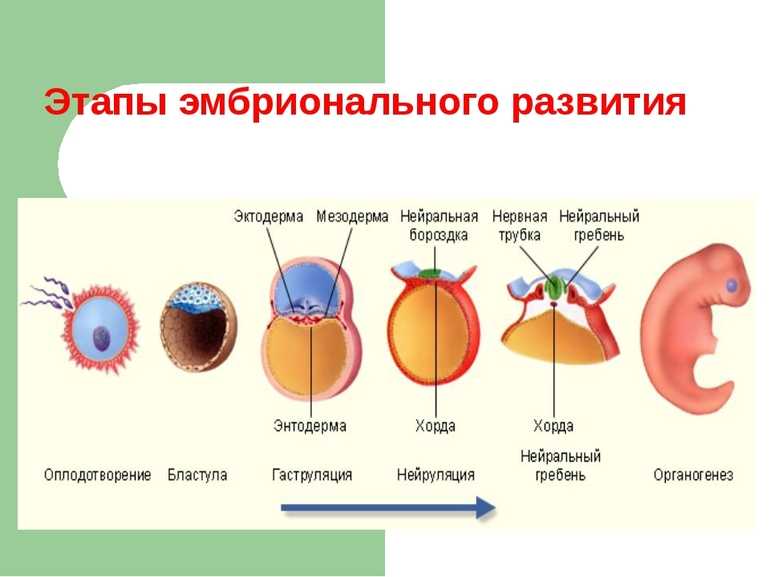


Микроспорогенез и образование пыльцевых зерен
Микроспорогенез протекает в мужской репродуктивной сфере – в андроцее. Каждый элемент андроцея – тычинка – состоит из тычиночной нити и пыльника. Внутри пыльника имеются пыльцевые гнезда, содержащие археспориальную ткань. В археспориальной ткани пыльников из каждой диплоидной материнской клетки микроспор в результате мейоза образуется четыре микроспоры. Каждая микроспора делится путем митоза и образуется двухклеточное пыльцевое зерно: одна клетка называется генеративной (в дальнейшем на ее основе образуются спермии), вторая – вегетативной (это остаток вегетативных клеток мужского гаметофита). Зрелое пыльцевое зерно покрыто двойной оболочкой: экзиной (внешней) и интиной (внутренней).
Опыление
Опыление – это процесс переноса пыльцы из пыльников на рыльце пестика. Существует два основных типа опыления: самоопыление и перекрестное опыление. При самоопылении происходит перенос пыльцы в пределах одного цветка, а при перекрестном опылении пыльца из пыльников одного цветка переносится на рыльце пестика другого цветка.
Для более точной характеристики опыления с точки зрения генетики используются специальные термины: клейстогамия (самоопыление в нераскрывшемся цветке), автогамия (самоопыление в раскрывшемся цветке), гейтоногамия (перенос пыльцы из пыльника одного цветка на рыльце пестика другого цветка в пределах одного растения), аллогамия (перенос пыльцы из пыльника цветка одного растения на рыльце пестика цветка другого растения). Таким образом, лишь аллогамия является собственно перекрестным опылением, а остальные типы опыления тождественны самоопылению.
Образование пыльцевой трубки и спермиев
В результате опыления пыльцевое зерно попадает на рыльце пестика и прорастает в пыльцевую трубку (мужской гаметофит). В пыльцевой трубке ядро генеративной клетки делится путем митоза, образуя два генеративных ядра. (У ряда растений деление генеративной клетки происходит еще в пыльниках). Каждое генеративное ядро с прилежащим слоем цитоплазмы называется спермием.
Двойное оплодотворение, образование зародыша и семени
Пыльцевая трубка внедряется в ткани столбика и растет в направлении завязи, при этом на вершине пыльцевой трубки находится вегетативное ядро, а за ним движутся оба спермия. Затем пыльцевая трубка проникает в семязачаток через пыльцевход – микропиле (у некоторых растений существует халазогамия – проникновение пыльцевой трубки в семязачаток через его ножку – халазу). Далее пыльцевая трубка лопается, ее содержимое изливается в зародышевый мешок, и происходит двойное оплодотворение. Один спермий сливается с ядром яйцеклетки, и образуется диплоидная зигота. Другой спермий сливается с центральным диплоидным ядром зародышевого мешка, и образуется триплоидное ядро эндосперма. Все остальные ядра зародышевого мешка и вегетативное ядро пыльцевого зерна – разрушаются.
Из диплоидной зиготы развивается зародыш, а из триплоидного центрального ядра – эндосперм. Происходит формирование семени, включающего зародыш, эндосперм и семенную кожуру, которая образуется из покровов семязачатка.
1.5. Жизненный цикл и размножение растений
В жизненном цикле растений чередуются две фазы: гаметофит
(гаплодиная) и спорофит (диплоидная). Спорофит размножается бесполым
путем: в мешковидных спорангиях в результате мейоза формируются
гаплоидные споры. Споры могут быть одинаковы (у равноспоровых растений), из них
прорастают обоеполые гаметофиты. У разноспоровых формируется два типа спор:
микро- и мегаспоры, при их прорастании формируются однополые гаметофиты –
мужские и женские. Соответственно спорангии называются микро- и мегаспорангии.
Спорангии располагаются на спороносных листовых структурах: микро- и
мегаспорофиллах. Спорофиллы располагаются поодиночке, либо собраны в
спороносные шишки – стробилы.
На гаметофите формируются гаметангии, в которых путем
митоза образуются гаметы. В мужских гаметангиях (антеридиях) формируются
подвижные жгутиковые сперматозоиды или неподвижные безжгутиковые спермии. В
женских гаметангиях (архегониях) формируются яйцеклетки. Архегонии имеют
бутылковидную форму: расширенное брюшко, в котором помещается одна или
несколько яйцеклеток, и шейка, в которой по мере созревания за счет разрушения
клеток образуется канал. Внутри архегония происходит оплодотворение и
формируется зигота, из нее вырастает диплоидный спорофит.
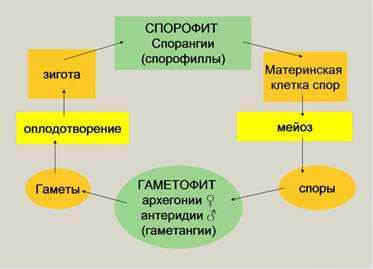
Рис. 11. Схема жизненного цикла растений
Жизненный цикл и размножение цветковых растений