Механизм действия
Действие иммунизирующих фунгицидов всегда является системным, в связи с чем, их также называют системными псевдофунгицидами. Проявлять свое влияние, находясь на поверхности растений, они не способны.
Термин «элиситор», который получил распространение в середине 80-х годов XX века, в смысловом переводе означает «вызывающий ответную реакцию». Собственно, именно к проявлению своеобразной ответной реакции и сводится механизм действия этой группы препаратов. Находясь внутри растительного организма, элиситоры:
- Стимулируют повышенное образование защитных метаболитов растения, повышая его иммунитет. Так, препараты меди, используясь для обработки горошка, приводили к усилению синтеза в тканях растений фитоалексина (защитного вещества) под названием пизатин. Это соединение обычно образуется в растении при заражении фитопатогенными грибами, а под влиянием профилактической обработки вырабатывается еще до инфицирования, препятствуя его наступлению.
- Усиливают синтез естественных соединений, при повышенной концентрации которых в тканях создаются условия, мало приемлемые для развития возбудителя. К примеру, триадимефон, помимо прочих компонентов действия, повышает в листьях растений концентрацию каротиноидов, ксантофилла и других факторов, в присутствии которых развитие патогенов замедляется.
- Приводят к образованию веществ, инактивирующих токсины возбудителей. Например, под действием производных фенилмочевины в плодах огурца и яблони повышалось количество фенолов; из них под действием процессов метаболизма возникали полифенолы, оказывающие ингибирующее влияние на пектолитические ферменты возбудителей парши яблони и оливковой плесени огурца.
Один и тот же препарат может проявлять сразу несколько механизмов действия. Процесс иммунизации запускается под влиянием либо самого средства, либо его метаболитов.
В химической защите с целью иммунизации применяют ряд фунгицидов, принадлежащих к различным группам. В некоторых случаях иммунизирующее действие лучше проявляется при сочетании нескольких ДВ с разными механизмами действия. К примеру, препарат Максим XL на основе флудиоксонила и мефеноксама был заявлен производителями как мощный элиситор.
Биохимические элиситоры устойчивости к заболеваниям
Известно, что штаммы Trichoderma образуют 3 типа веществ, которые индуцируют резистентность растений. Это белки с ферментативной активностью или с какой-нибудь другой активностью, продукты генов Avr (антивирулентные белки), олигосахариды и низкомолекулярные вещества, которые выделяются из клеточных стенок грибов и растений в результате воздействия на них ферментов Trichoderma.
Даже до того, как было установлено, что штаммы Trichoderma способны индуцировать резистентность растений, было известно, что ксиланаза 22 kDa, которую секретируют многие виды, индуцирует синтез этилена и защитные реакции растений. Интересно, что этот небольшой белок быстро переносится в сосудистой системе растения табака при введении через разрез листового черенка. Белок индуцировал локальную резистентность и некроз, для развития СРР белок должен распространиться по всему растению.
Недавно установлено, что штамм T. virens выделял серию белков и пептидов, которые индуцировали биосинтез терпеноидных фитоалексинов и пероксидазы в растениях хлопчатника. Показано, что 6 белков и пептидов обладали элиситорной активностью. Один из них обладал кросс-иммунореактивностью к ксиланазе, индуцирующей синтез этилена в растениях, а другой размером 18 kDa в области NH2 конца был гомологичен сериновой протеиназе Fusarium sporotrichiodes (Harman et al.,2004).
Фитопатологическая оценка развития патогенов
Для измереная концентрации перекиси водорода использовали методику в применением красителя ксиленолового оранжевого (Bindschedler et al., 2001). Реагент содержал 0,074% соли Мора в 5,81% растворе серной кислоты и 0,009% раствор ксиленолового оранжевого в 1,82 % растворе сорбита (в соотношении 1:100). Для определения к 500 мкл реагента добавляли 50 мкл супернатанта образца, инкубировали при комнатной температуре 45 мин. Перед измерением образцы центрифугировали при 16000 об/мин в течение 5 мин, затем измеряли оптическую плотность образовавшегося комплекса на спектрофотометре Biospeck-Mini («Shimadzu», Япония) при длине волны 560 нм. Концентрацию Н202 определяли по предварительно построенной калибровочной кривой.
Растительный материал (листья пшеницы или часть клубня картофеля) растирали с жидким азотом и извлекали растворимые белки, используя сахарозный буфер и фенол в соответствие с рекомендациями (Vincent et al, 2006). Пробы центрифугировали 30 мин при 4400 об/мин. Для осаждения белка к супернатанту приливали ледяной раствор ацетата аммония в спирте в соотношении 1:4. Белки выдерживали 1 ч при -20С. Осажденный белок промывали три раза охлажденным спиртом и хранили при -70 С. Белок растворяли в буфере, содержавшем 8М мочевину, 2М тиомочевину, 1% CHAPS, 30 мМ ДТТ, 20 мМ трис основной, 0.3% раствор амфолитов, рН 3-Ю.
Для количественного определения белка к 20 мкл исследуемого образца приливали 1 мл реактива Брэдфорд (10 мг Кумасси G-250 в 5 мл этанола + 10 мл ортофосфорной кислоты и довести до 100 мо дистиллированной водой). Оптическую плотность полученного раствора измеряли на спектрофотометре Biospeck-Mini («Shimadzu», Япония) при 595 нм через 5 мин, но не позднее 1 часа. Концентрацию белка определяли по калибровочному графику. Для построения калибровочной кривой использовали растворы бычьего сывороточного альбумина с известной концентрацией от 1 до 0,1 мг/мл.
Изоэлектрофокусирование белков проводили на приборе Protean IEF (“Biorad”, США). Для разделения белков в первом направлении (по изоэлектрической точке) использовали готовые 7-сантиметровые стрипы (“Biorad”, США), диапазон рН 3-Ю. Перед фокусированием проводили пассивную регидратацию в течение 12 часов при 20С. Фокусирование проводили при напряжении 4000 В (20000 Вч) в течение 22 часов, затем поддерживали напряжение 500 В до окончания процесса. После изоэлектрофокусирования стрипы выдерживали по 15 минут последовательно в растворах 2%-ного дитиотреитола и 2.5%-ного йодацетамида в буферных растворах с 25%-ным глицерином, затем промывали в 0.025М трис-глициновом буфере, рН 8.3.
Для разделения белков по молекулярной массе проводили SDS-электрофорез в 12%-ном ПААГ по Лэммли. Стрип и маркерные белки на фильтровальной бумаге помещали на полиакриламидный гель и проливали 1%-ной агарозой на трис-глициновом буферном растворе. Электрофорез вели при напряжении 90-120В с помощью источника питания “Эльф” (“ДНК-технология”, Россия) в камере для вертикального электрофореза (“Biorad”, США), гели стабилизировали в 50%-ном этаноле в течение 10 минут, затем окрашивали 0.1%-ным раствором Кумасси G-250 в течение 10 часов при комнатной температуре. Гели отмывали от красителя в растворе, содержащем 8% ледяной уксусной кислоты и 25% этанола. 2.3.4. Выделение и очистка РНК из растений
Для выделения РНК из растений использовали видоизмененный метод Chomezynski (1997). Для этого листья (корни) растений растирали в жидком азоте до гомогенного состояния. К полученному объему добавляли 1 мл тризола. Затем добавляли равное количество водонасыщенного (рН 5,0) фенола и хлороформа, интенсивно перемешивали на встряхивателе типа 358S («Elpan», Польша) и центрифугировали при 10000 об/мин в течение 20 минут при 4С. Водную фазу переносили в чистую пробирку с равным объемом водонасыщенного фенола и хлороформа, вновь тщательно перемешивали и центрифугировали при тех же условиях. На последнем этапе выделения добавляли изопропанол и оставляли при -20С минимум на один час для формирования осадка РНК. Перед использованием РНК осаждали центрифугированием, промывали 70% этанолом и растворяли в минимальном количестве дистиллированной воды, обработанной DEPC. Для более тщательной очистки от примесей ДНК осадок РНК осаждали 8М LiCl в течение ночи. Все процедуры с РНК проводили при 4С.

Статья составлена с использованием следующих материалов:
Бушковская Л.М., Марчук Т.Л. и др. Эффективность бактофита на лекарственных культурах. Тезисы докладов Всероссийского съезда по защите растений «Защита растений в условиях реформирования агропромышленного комплекса: экономика, эффективность, экологичность», Санкт-Петербург, декабрь 1995 г., с.293-294
3.Голышин Н. М. Фунгициды. — М.: Колос, 1993. -319 с.: ил.
4.Груздев Г.С. Химическая защита растений. Под редакцией Г.С. Груздева — 3-е изд., перераб. и доп. — М.: Агропромиздат, 1987. — 415 с.: ил.
5.
Зинченко В.А. Химическая защита растений: средства, технология и экологическая безопасность. – М.: «КолосС», 2012. – 127 с.
6.
Штерншис М.В., Томилова О.Г., Андреева И.В. Биотехнология в защите растений: Учеб.пособие/ Министерство сельского хозяйства РФ. Новосиб.гос.аграр.ун-т. – Новосибирск, 2001. – 156 с.
Источники из сети интернет:
7.
http://www.syngenta.com
Изображения (переработаны):
8.
Fusarium wilt Fusarium oxysporum f.sp. lycopersici, by R.W. Samson, Purdue University, Bugwood.org, по лицензии CC BY-NC
Свернуть
Список всех источников
Сравнительная оценка транскрипционной активности генов защитных белков в растениях пшеницы с различной устойчивостью к Septoria nodorum
Как показали исследования, в ответ на инфицирование возбудителем септориоза через 24 ч после инокуляции S. nоdоrum происходит усиление экспрессии гена оксалатоксидазы (рисунок 12 — 2) и повышение активности фермента (рисунок 11 — 2). В последующем (через 48 и 72 ч после инокуляции) экспрессия гена оксалатоксидазы (рисунок 12 — II, III, 2) и, соответственно, активность фермента (рисунок 11 — II, III, 2) снижаются.
ХОС в зависимости от СА проявляли различное стимулирующее действие на экспрессию гена оксалатоксидазы, как в неинфицированных, так и инфицированных листьях (рисунок 12). Так под воздействием ХОС со СА 65% уровень экспрессии гена оксалатоксидазы и активность фермента превосходили аналогичный показатель инфицированных листьев пшеницы, обработанных ХОС со СА 30%, на протяжении всего опыта (рисунок 11, 12 — 4, 6). Причем, максимальные различия в индукции экспрессии гена оксалатоксидазы и активности фермента между ХОС со СА 65% и СА 30% наиболее контрастно проявлялись в неинфицированных растениях (рисунок 11, 12 — 3, 5).
Повышение активности оксалатоксидазы и усиление продукции перекиси водорода при обработке ХОС со СА 65% показано ранее при инфицировании каллусов пшеницы возбудителем твердой головни (Davoine et al., 2001)
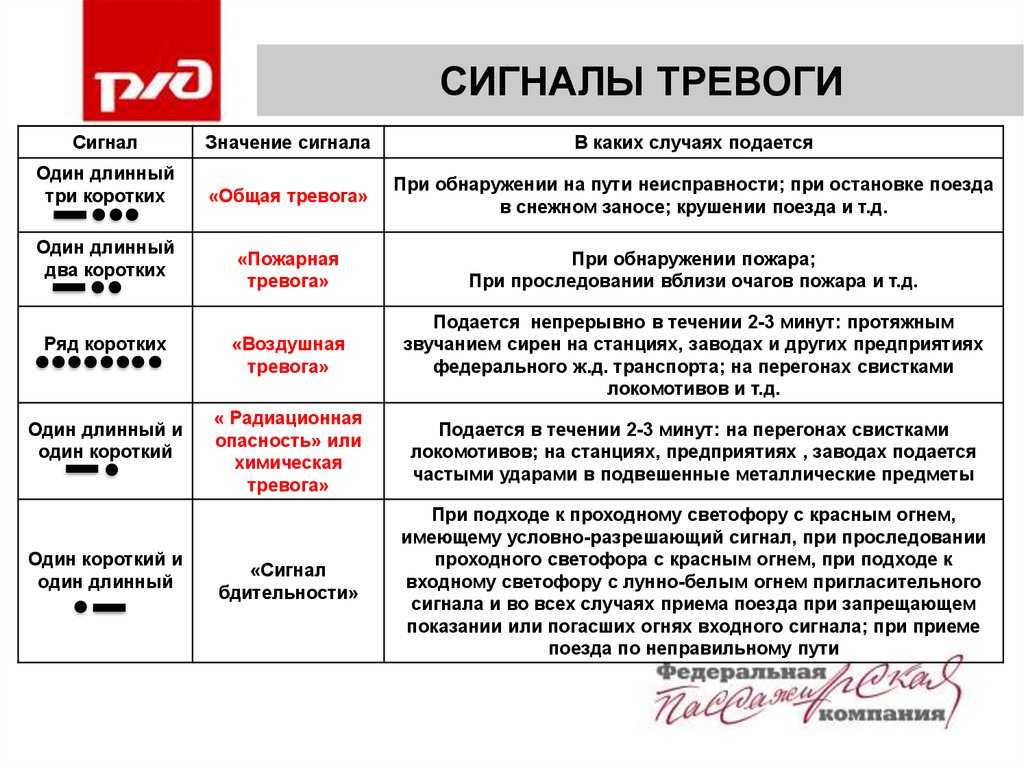
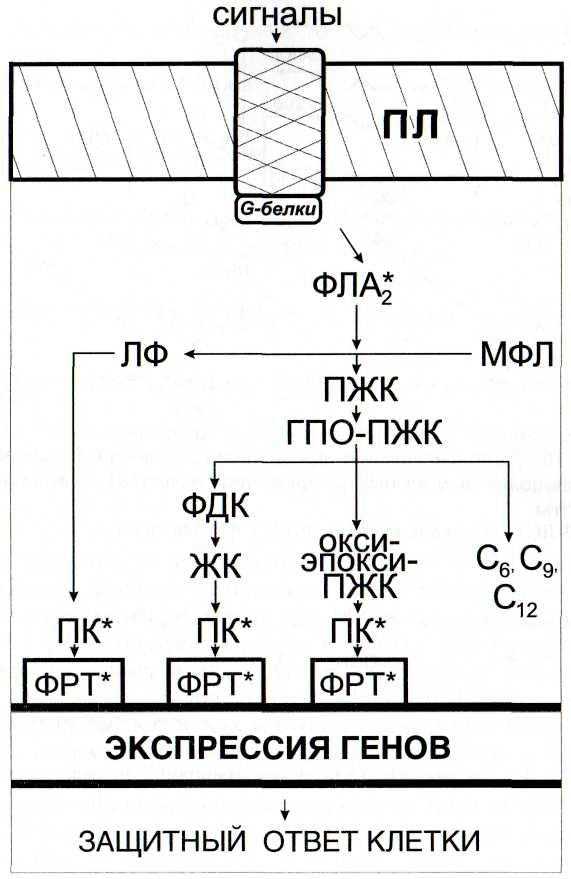
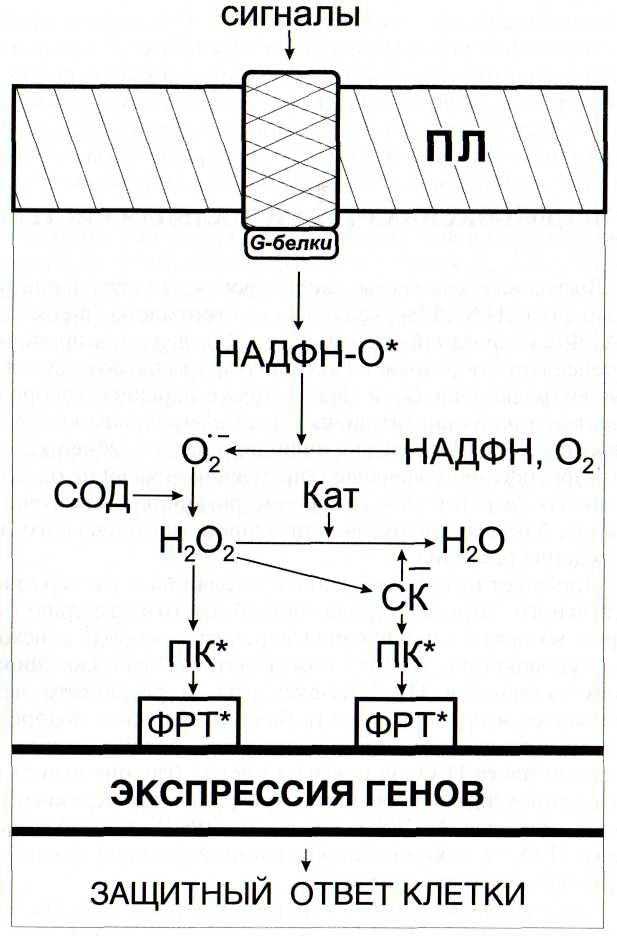
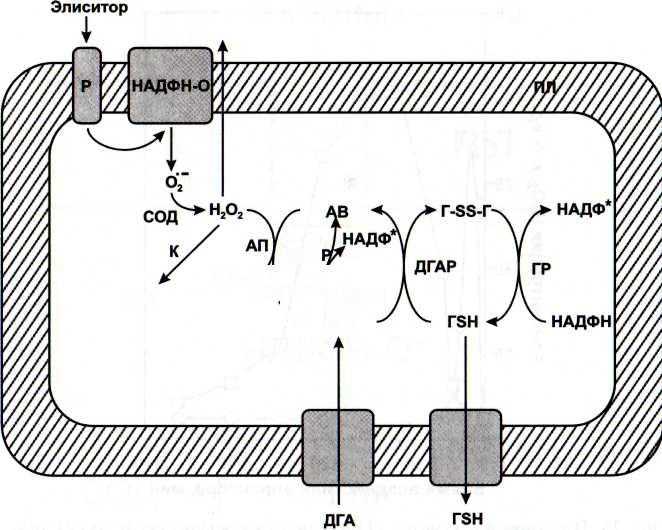
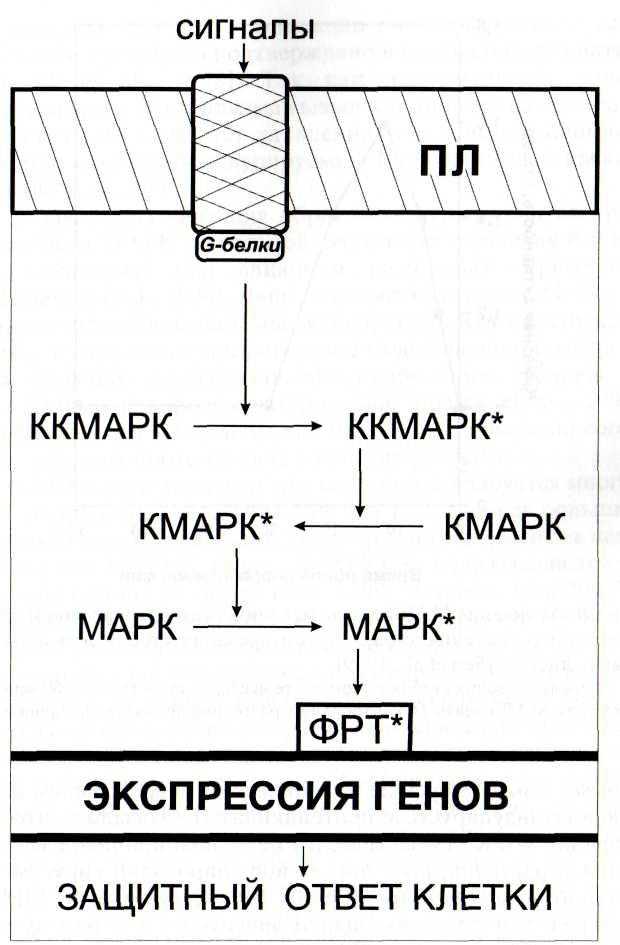
В настоящее время известно, что активные формы кислорода, в том числе, перекись водорода, являются важной составляющей ответных реакций на внедрение патогенов. Защитное действие Н202 обусловлено как прямым биоцидным действием на патоген, так и ее участием в сигнальных системах растительных клеток
В связи с открытием сигнальной и защитной роли активных форм кислорода большое внимание уделяется оксидоредуктазам, регулирующим их уровень в клетке (Francoz et al, 2015). Среди них особый интерес представляют пероксидазы, активность которых коррелирует с развитием устойчивости растений к патогенам (Kawano et al., 2003)
Показано, что анионная пероксидаза принимает непосредственное участие в синтезе лигнина, ограничивающего проникновение в ткани растения инфекционных структур гриба (Минибаева, Гордон, 2003). 16
Исследования показали, что в ответ на инфицирование S. nоdоrum происходит усиление тарнскрипционной активности гена анионной пероксидазы (рисунок 13 — 2), и повышается активность фермента (рисунок 13 — 2). Причем, ХОС со СА 30% оказывали незначительный стимулирующий эффект на экспрессию гена анионной пероксидазы в неинфицированных листьях пшеницы (рисунок 14 — 3). Однако при инфицировании S. nоdоrum стимулирующее действие ХОС со СА 30% заметно повышалось (рисунок 14 — 4), что положительно отражалось на активности фермента (рисунок 13 — 4). Так через 48 ч после инокуляции уровень экспрессии гена анионной пероксидазы превышал 400 350 300 250 200 150 100 контрольный вариант в 2 раза, а через 72 ч после инокуляции — в 2,5 раза (рисунок 14 — 4).
Индуцирующий эффект ХОС со СА 65% на экспрессию гена анионной пероксидазы оказался более значительным и продолжительным как в неинфицированных (рисунок 14 — 5), так и инфицированных листьях (рисунок 14 — 6). Так уже через 24 ч после инокуляции у пшеницы, обработанной ХОС со СА 65%, уровень экспрессии гена анионной пероксидазы превышал контрольный вариант в 2,6 раза и оставался высоким на протяжении 72 ч (рисунок 14 — 6), что отражалось на активности фермента (рисунок 13 — 6). Сходный эффект ХОС со СА 65% на активацию пероксидазы был обнаружен при инфицировании растений пшеницы возбудителем корневой гнили Bipоlаns sоrоhmаnа (Huckelboven, Kogel, 2003).
Таким образом, ХОС со СА 65% оказывают более значительное индуцирующее действие на экспрессию генов оксалатоксидазы и анионной пероксидазы по сравнению с ХОС со СА 30%. Возможно, более высокая чувствительность генов оксалатоксидазы и анионной пероксидазы к ХОС со СА 65% объясняется тем, что данные ферменты участвуют в регуляции уровня Н202 на самых ранних этапах инфекционного процесса. Оксалатоксидаза окисляет щавелевую кислоту и оксалаты с образованием перекиси водорода, которую пероксидаза использует в реакциях лигнификации. Учитывая, что хитин входит в состав клеточной стенки грибов, то в процессе его разрушения хитиназами, на начальных этапах патогенеза образуются хитиновые фрагменты с высокой степенью ацетилирования, которые вовлекаются в регуляцию активности ферментов метаболизма АФК. Такой механизм индукции защитного ответа ХОС мог сформироваться в процессе коэволюции системы «растение — грибной патоген».
Влияние Trichoderma на потребление питательных веществ растениями
Виды Trichoderma увеличивали потребление и концентрацию разнообразных элементов (меди, фосфора, железа, марганца и натрия) корнями растений в гидропонной культуре даже в неблагоприятных условиях (Yedidia et al., 2001). Увеличение потребления свидетельствует о положительном влиянии Trichoderma на интенсивность потребления элементов растениями. Кукуруза реагировала на повышение усвоения азотных удобрений тем, что листья растений становились более зелеными, чем в контроле. Урожайность достигала максимальных показателей и выходила на плато. Значения максимальной урожайности специфичны для каждого генотипа в соответствующих условиях. Однако было показано, что культуры, выращенные из семян, протравленных Т-22, дают максимальные урожаи, хотя под них вносили на 40% меньше азотных удобрений, чем в контроле (Harman et al., 2000). При дополнительной азотной подкормке урожай может превысить значения плато для данной культуры. Результаты показывают, что Т-22 увеличивает эффективность азотного удобрения на кукурузе. Данное обстоятельство позволило сократить нитратное загрязнение почвы и поверхностных вод, что является серьезным побочным отрицательным следствием крупномасштабного производства кукурузы. Как показывают анализы, кроме положительного влияния на усвоение азота, Т-22 значительно увеличивал потребление многих элементов, таких, как мышьяк, кобальт, кадмий, хром, никель, свинец, ванадий, магний, марганец, медь, бор, цинк, алюминий и натрий. В полевых опытах эффект значительно снижался (Harman, 2004).
Наконец, известно, что Т-22 – и, возможно, другие виды Trichoderma – способны переводить в растворимые формы различные питательные для растений элементы, такие, как минеральные фосфаты, Fe3+, Cu2+, Mn4+, Zn0. Эти элементы часто недоступны в почве для растений. Т-22 снижает содержание окисленных ионов металлов, переводит их в подвижные формы, выделяет сидерофоры, хелатирующие ионы. Исследована способность стимуляции роста растения и биоконтроля гриба Trichoderma harzianum Rifai 1295-22 (Т-22) путем растворения in vitro некоторых нерастворимых или недостаточно растворимых минералов (Altomare et al., 1999). Существует три возможных механизма: подкисление среды, продуцирование хелатных метаболитов и окислительная активность. Т-22 был способен к растворению MnO2, металлического цинка и неорганического фосфата (главным образом, фосфата кальция) в жидкой среде, содержащей сахарозно-дрожжевой экстракт. Подкисление не было главным механизмом растворения, так как pH культур никогда не падал ниже 5.0, и в культурах, содержащих MnO2, pH повышался от 6.8 до 7.4. Органические кислоты не были обнаружены. Fe2O3, MnO2, Zn, и неорганический фосфат были растворены. Эта деятельность может объяснять, по крайней мере, частично, способность T-22 к увеличению роста растения. Растворение металлической окиси происходит с участием хелатиона и процесса окисления. Оба из этих механизмов также играют роль в биоконтроле патогенных растений при разнообразных состояниях окружающей среды.
Радриш с соавторами (Rudresh et al., 2005) исследовали эффект комбинированной инокуляции Rhizobium, фосфат растворяющих Bacillus megaterium sub sp phospaticum и биоконтроль Trichoderma в росте, потреблении азота и урожая нута в теплице и в полевых условиях. Комбинированная инокуляция этих трех организмов привела к повышению прорастания, усваивания азота (120 мг/раст.), фосфора (6 мг/раст.), высоты растений, числа ответвлений, образования узелков, урожая, и общей биомассы по сравнению с каждой индивидуальной инокуляцией и не инокулированным контролем.
Улучшение корневой системы и роста растений
Виды Trichoderma и другие полезные колонизирующие корни микроорганизмы улучшают рост растений и увеличивают урожайность культур, но долговременное улучшение могут вызывать только ризосферокомпетентные штаммы. В тепличном растениеводстве штамм Т-22 T. harzianum широко используется для защиты от болезней растений вместо химических фунгицидов, поскольку он безопасен для работников. Эффект биоконтрольного агента сохраняется дольше, чем эффект пестицидов, препарат дешевле пестицидов, препарат, в отличие от пестицидов, улучшал состояние корневой системы растений (Harman et al., 2000).
Продукты Avr генов
Белковые продукты Avr генов идентифицированы у большого количества фитопатогенных грибов и бактерий. Они расоспецифичные и индуцируют гиперчувствительность или другие защитные реакции в культиварах растений, которые содержали соответствующие гены резистентности. С помощью протеомного анализа штамма Т-22 идентифицированы белки, гомологичные Avr4 и Avr9 Cladosporium fulvum. Штамм Р1 T. atroviride также образовывал подобные белки, олигосахариды и низкомолекулярные соединения. Получены мутанты Trichoderma, экспрессирующие сигнальные системы, такие, как зеленый флюоресцентный белок или глюкозоксидаза под контролем промоторов, ассоциированных с биоконтролем. Это позволило выделить и охарактеризовать биологически активные молекулы, которые освобождаются из клеточных стенок фитопатогенных грибов или из растений под воздействием гидролаз Trichoderma. Эти молекулы, образующиеся в результате многообразных взаимодействий Trichoderma с патогенными грибами и с корнями растений в природе, действуют как индукторы экспрессии антагонистического каскада генов Trichoderma и как элиситоры защитных механизмов растений (Kubicek et al., 2001).
Похожие патенты RU2080788C1
| название | год | авторы | номер документа |
|---|---|---|---|
| ИНДУКТОР УСТОЙЧИВОСТИ КАРТОФЕЛЯ К ФИТОПАТОГЕНАМ | 1991 |
|
RU2013037C1 |
| ЗАЩИТА РАСТЕНИЙ С ИСПОЛЬЗОВАНИЕМ РЫБЬЕГО ЖИРА | 1995 | RU2170509C2 | |
| Штамм бактерий Bacillus subtilis Б-94 ВИЗР для защиты картофеля от болезней при вегетации и хранении, повышения урожайности и улучшения качества клубней | 2019 |
|
RU2729576C1 |
| Средство для предпосевной обработки клубней картофеля | 2018 |
|
RU2694202C1 |
| КОМПОЗИЦИЯ НА ОСНОВЕ ВОДНЫХ РАСТВОРОВ ХИТОЗАНА, ОБЛАДАЮЩАЯ БИОЛОГИЧЕСКОЙ АКТИВНОСТЬЮ | 1997 |
|
RU2127056C1 |
| Способ изменения фенотипа и устойчивости растений | 2021 |
|
RU2780853C1 |
| СТИМУЛЯТОР РОСТА РАСТЕНИЙ | 1994 |
|
RU2076598C1 |
| БЕЛКИ, ИНДУЦИРУЮЩИЕ МНОЖЕСТВЕННУЮ УСТОЙЧИВОСТЬ РАСТЕНИЙ К ФИТОПАТОГЕНАМ И ВРЕДИТЕЛЯМ | 2004 |
|
RU2333220C2 |
| КОМПОЗИЦИЯ ДЛЯ ИММУНИЗАЦИИ РАСТЕНИЙ ОТ РАЗЛИЧНЫХ ФИТОПАТОГЕНОВ | 1994 | RU2075933C1 | |
| СПОСОБ ПОЛУЧЕНИЯ ФОРМ КАРТОФЕЛЯ in vitro, УСТОЙЧИВЫХ К ВОЗБУДИТЕЛЯМ ФИТОФТОРОЗА И АЛЬТЕРНАРИОЗА | 2013 |
|
RU2524424C1 |
Особенности иммунизаторов биологического происхождения
Действие иммунизаторов биологического происхождения часто сводится не столько к выработке защитных факторов, сколько к конкуренции с патогенами за приоритет заражения культур. Для этого применяются авирулентные или гиповирулентные штаммы возбудителей, которые, если бы они находились в активной форме, были бы потенциально способны вызвать заражение. Они выделяются из природных объектов или получаются искусственным способом, при помощи воздействия на них физико-химическими и другими факторами. Например, безвредные штаммы Rhizocionia solani защищают растения от инфицирования вирулентными формами этих же микроорганизмов, развиваясь внутри них, но не принося им вреда.
Эффективность таких средств довольно высока. Так, при воздействии на растения табака ослабленными возбудителями пероноспороза в них происходило накопление полимеров и фитоалексинов, и возрастала активность защитных ферментов хитиназы и пероксидазы, что приводило к повышению устойчивости. Аналогичным образом, иммунизация штаммами Fusarium охуjporum в полевых условиях была сопоставима с применением беномила. В результате обработки томаты и баклажаны оказались защищены от фузариозного увядания и других заболеваний.
Некоторые средства биологической защиты не заражают растения, а действуют другими способами. Например, ряд биофунгицидов воздействуют на состояние ризосферы, изменяют кислотность и другие показатели почвы, не допускают развития в ней плесневых грибов и других вредных микробов, улучшают усвоение растениями питательных веществ. Это может сочетаться со стимуляцией выработки в культуре естественных антибиотиков. Подобное комплексное действие, в частности, наблюдается у бактерий Bacillus subtilis, штамм ИПМ 215.
Влияние колонизации корней на метаболизм растения
В некоторых работах показано, что колонизация корней видами Trichoderma приводит к увеличению уровня защитных ферментов растений, в том числе пероксидаз, хитиназ, β-1,3-глюканаз и фермента липоксигеназного пути гидроксипероксид лиазы (Howell et al., 2000). Так, внесение штамма Т-203 T. asperellum под огурцы приводило к постоянному увеличению синтеза фенилаланин-аммиаклиазы в проростках и корнях, но через 2 дня эффект исчезал. Однако если на листья наносили инокулят патогена Pseudomonas syringae pv. lachrymans, экспрессия генов защитных белков продолжалась длительное время.