
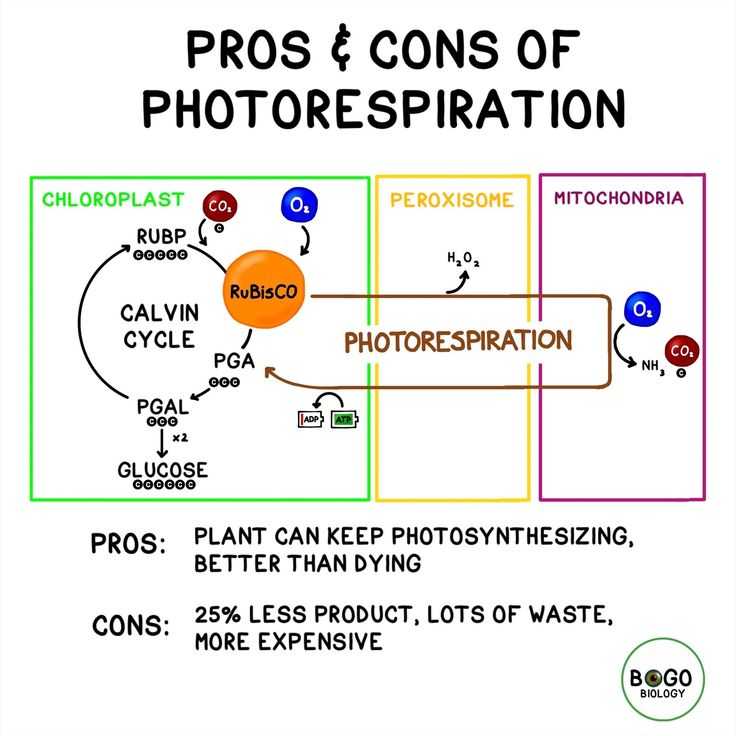
ВВЕДЕНИЕ
Классические представления о донорно-акцепторной системе (ДАС) растений сложились
в прошлом столетии как результат исследования взаимодействия органов, производящих
и потребляющих ассимиляты . Однако одни и те же части растения, например, листья, могут одновременно образовывать
и использовать ассимиляты, а корни и другие органы – импортировать и экспортировать
продукты фотосинтеза []. Концепция донорно-акцепторных отношений (ДАО) во многом способствовала формированию
представлений о растении как целостной системе. Особую роль в этом сыграли работы
А.Т. Мокроносова , рассматривавшего растение как систему доноров и акцепторов, взаимодействие которых
осуществляется благодаря канализированным системам транспорта веществ и передачи сигналов.
В процессе реализации морфогенетической программы роста и развития в растении возникают
все новые донорные листья и сменяют друг друга акцепторы, что свидетельствует о динамичности
донорно-акцепторных связей. Благодаря своей универсальности, донорно-акцепторная концепция
логично вписалась в общую теорию продукционного процесса и стала ее неотъемлемой частью.
Согласно А.Т. Мокроносову [], фотосинтезу здесь отводится место важнейшей функции растительного организма, ему
подчиненного и им управляемого.
Развитие представлений о ДАС способствовало разработке идеи оптимального морфофизиологического
типа растения для создания высокопродуктивных посевов сельскохозяйственных культур
[]. По современным представлениям продукционный процесс является интегральной функцией
растений, основу которой составляют фотосинтез, дыхание и рост. Важную роль в формировании
продуктивности играют процессы распределения углерода между органами, которые контролируются
системой прямых и обратных связей, образующих единую донорно-акцепторную систему растений.
Основные регуляторные функции в ДАС отводятся фитогормонам, механизмам транспорта
ассимилятов и аттрагирующему действию акцептора. А.Т. Мокроносов не исключал также
способности донора влиять на загрузку, транспорт и распределение ассимилятов [].
Реализация генетически запрограммированной программы роста и развития растений в значительной
степени зависит от условий внешней среды. В этом отношении интерес представляет изучение
ДАС культурных растений на северной границе возделывания, где их продуктивность ограничивается
коротким периодом вегетации, недостатком тепла, перепадами температуры, низким естественным
плодородием почв.
Чем отпереляется скорость пережвижения веществ по флоэме?
7 декабря, 2010
Скорость передвижения веществ по флоэме определяли, наблюдая за быстротой распространения меченых соединений. Оказалось, что скорость передвижения в ситовидных трубках достаточно высока и составляет в среднем 50—100 см/ч. У разных групп растений этот показатель может несколько варьировать. У одного и того же растения различные органические вещества могут передвигаться с разной скоростью. Значительное влияние на скорость передвижения оказывают условия внешней среды. В отличие от перемещения по ксилеме на транспорт веществ по флоэме влияют все факторы, изменяющие напряженность процессов обмена веществ. Передвижение по флоэме зависит от температуры. Это можно проследить на опытах, в которых пластинки листа погружали в раствор сахарозы, а черешки заключали в специальную муфту и подвергали воздействию различных температур. Оказалось, что оптимальная температура колеблется между 20 и 30°С. Дальнейшее повышение температуры уже тормозит отток ассимилятов из пластинки листа. Отношение к резкому охлаждению флоэмы у разных растений неодинаково. У южных растений (фасоль) полностью приостанавливается транспорт при температуре 1—2°С, тогда как у сахарной свеклы подобное охлаждение лишь замедляет передвижение. Условия минерального питания оказывают заметное влияние на транспорт веществ по флоэме. Особенно много исследований посвящено влиянию бора. Известно, что под влиянием бора скорость передвижения сахарозы заметно возрастает. Скорей всего, это связано с образованием комплексных соединений бора с углеводами. Скорость передвижения ассимилятов увеличивается и под влиянием фосфора. Фосфорилированные формы сахаров передвигаются быстрее. Скорость передвижения меняется под влиянием калия. В последнее время появилась гипотеза, согласно которой калий поддерживает мембранный потенциал в ситовидных пластинках и тем самым способствует передвижению ассимилятов. Передвижение веществ по флоэме тормозится в присутствии всех метаболических ингибиторов, таких, как азид натрия, йодацетат, динитрофенол и др. Транспорт по флоэме ускоряется при добавлении АТФ. Все это бесспорно указывает на тесную связь между передвижением веществ по флоэме и метаболизмом.
Изучение механизма аттрагирующего эффекта цитокининов в изолированных листьях на уровне структуры флоэмы
Поскольку не исключено было участие флоэмы в реализации аттрагирующего эффекта цитокинина в изолированных листьях (Miiller, Leopold 1966 a; Nakata, Leopold, 1967; Leonard, King, 1968; Penot et al., 1981; Gersani, Kende, 1982), мы изучили действие БАП на структурную организацию этой ткани у листьев тыквы и бобов, имевших открытую и закрытую терминальную флоэму соответственно. Систему жилкования изучали у листьев интактных растений тыквы и бобов, а также у половинок изолированного листа, одна из которых была опрыснута водой, а другая БАП на тех фазах развития листа, на которых проявлялся аттрагирующий эффект цитокининов, т.е. на достигших 50, 100 % Smax и стареющих листьях. У обоих растений жилкование было перистым (разновидность сетчатого, с одной главной жилкой). Мелкими жилками (к которым принято относить жилки двух последних порядков ветвления) у них были жилки 5-6 порядков. У тыквы ареолы были небольшими, средняя площадь одной ареолы в растущих листьях была равна 0.03 мм2, в зрелых и стареющих — 0.10. Свободные окончания внутри ареол практически отсутствовали. Лишь 13-15% ареол у растущих и около 25% у зрелых и стареющих листьев содержали по одному неветвящемуся окончанию (табл. 27). Мелкие жилки в листьях бобов образовывали более сложную анастамозную сеть со свободными простыми или разветвленными окончаниями внутри ареол. В листьях этого растения ареолы были достаточно крупными.
Средняя площадь одной ареолы составляла в растущих листьях около 0.35 мм2, в зрелых и стареющих — около 0.40. Чуть менее половины ареол не содержали свободных окончаний, большая часть (около 40%) ареол І имела по одному неразветвл енному окончанию, около 10% — по два простых неразветвленных окончания, около 4% — по одному разветвленному окончанию и в 0.1-0.2% случаев в ареоле содержалось либо по 3 простых, либо по 2 разветвленных окончания проводящей системы (табл. 28). Результаты количественной оценки жилкования в опытном и контрольных ва риантах показали, что обработка половинки листа БАП (опыт), как и просто его срезание (контроль) ни у тыквы, ни у бобов не изменяли жилкования.
На всех изученных стадиях развития листа оставались постоянными число порядков ветвления, число свободных окончаний в ареоле,площадь одной ареолы и общее число ареол в одной половинке листа (табл. 27, 28). Хотя клеточный состав мелких жилок и ультраструктурная организация их отдельных элементов в норме у листьев тыквы и бобов были изучены ранее другими авторами (Ziegler, 1965; Turgeon et al„ 1975; Гамалей, 1990 и др.), было необходимо провести их предварительное изучение как контрольного варианта. В состав мелких жилок листа тыквы и бобов входило несколько типов клеток: трахеальные (а у тыквы и паренхимные) элементы ксилемы, ситовидные и паренхим-ные элементы флоэмы. Последние подразделялись на две разновидности: собственно клетки флоэмной паренхимы и клетки-спутницы, участвующие в загрузке ассимиля-тов из мезофилла во флоэму и встречающиеся только в мелких жилках (рис. 38). На поперечных срезах листья тыквы имели симметричные биколлатеральные мелкие жилки с двумя флоэмными полюсами (рис. 38 а). Однако, как известно (Schmitz et al., 1987; Гамалей, 1990), в жилках подобного типа адаксиальный полюс изолирован от мезофилла и потому не принимает участия в загрузке флоэмы. Загрузка идет на противоположном — абаксиальном — полюсе благодаря наличию в нем специализированных «загрузочных узлов» — симпластически связанных с мезофиллом комплексов клетка-спутница — ситовидный элемент. Поэтому именно этот участок терминальной флоэмы и подвергали электронно-микроскопическому контролю. Проводящие пучки на поперечных срезах листьев бобов были коллатеральными (то есть имели лишь один флоэмиый полюс) и симметричными. Флоэма состояла из двух блоков. В состав каждого из них входили один — два ситовидных элемента и две спутницы. Блоки располагались друг над другом в направлении от палисадного к губчатому мезофиллу и были разделены паренхимными клетками флоэмы (рис. 38 б).
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проведены в подзоне средней тайги (61°67′ с.ш., 50°76′ в.д.). Климат
района умеренно-континентальный с продолжительной холодной многоснежной зимой и умеренно-теплым
летом []. Поздние весенние заморозки возможны вплоть до середины июня, а ранние осенние –
нередки в начале августа. Сумма средних суточных температур выше 10°С варьирует в
пределах 1000–1200°С. Средняя многолетняя температура самого теплого месяца июля равна
16.6°С. Обеспеченность растений влагой удовлетворительная, за вегетационный период
выпадает в среднем 250 мм осадков. Характерной особенностью региона, расположенного
в первой световой зоне, является длинный день. В июне продолжительность светового
дня на широте г. Сыктывкара составляет 19 ч 30 мин.
В опытах использовали два вида картофеля: Solanum tuberosum L. (сорт Приекульский ранний и среднепоздний cорт Идеал) и S. andigenum Juz. et Buk. Растения выращивали на опытном участке с типичной подзолистой среднеокультуренной
почвой. Высадку клубней проводили в первой декаде июня на площади питания 0.3 × 0.9
м. Всходы появлялись в конце июня, начало клубнеобразования отмечали во второй декаде
июля. Рост растений характеризовали по накоплению биомассы и листовой поверхности.
Площадь листа каждого яруса определяли методом отпечатков листовых пластинок на хлоркобальтовой
бумаге.
Обработку растений 20% раствором суперфосфата с добавлением 0.05% 2,4-дихлорфеноксиуксусной
кислоты (2,4-Д) для ускорения старения и отмирания ботвы проводили в период интенсивного
налива клубней, за месяц до уборки урожая. Воздействие коротким днем (11 ч) осуществляли
в течение 20 сут после появления зачатков бутонов.
Лист определенного яруса растений сорта Приекульский ранний экспонировали в атмосфере
с 14СО2 в течение 20 мин. Эксперименты с меткой проводили в первой половине дня при солнечной
погоде, используя для этого полевой прибор, сходный с описанным в работе А.Т. Мокроносова
[]. Меченую углекислоту генерировали из Ва14СО3 при добавлении 0.1 н раствора HCl. Концентрация 14СО2 в газовой смеси составляла 1%, удельная радиоактивность 9.3 × 105 Бк/л. Распределение метки по органам растений изучали сразу, через сутки и спустя
неделю после экспозиции в атмосфере с 14СО2 индивидуальных листьев главного побега (пятый, десятый и пятнадцатый лист, считая
снизу).
Влияние внутренних факторов на поступление солей в растительный организм
2 декабря, 2010
Зависимость поступления солей от интенсивности дыхания является установленным фактом. При замене кислорода азотом не только прекращается поступление, но наблюдается выделение питательных ионов из корня. Ингибиторы процесса дыхания (в частности, цианистый калий) резко тормозят поступление солей. Процесс дыхания может оказывать влияние на поступление солей в нескольких направлениях. Так, в процессе дыхания выделяющийся углекислый газ в водной среде диссоциирует на ионы Н+ и НС03—. Адсорбируясь на поверхности корня, эти ионы служат обменным фондом для поступающих катионов и анионов. В процессе переноса ионов через мембрану участвуют специфические белки-переносчики, синтез которых находится в зависимости от интенсивности дыхательного процесса. Наконец, энергия, выделяемая в процессе дыхания, непосредственно используется для поступления солей (активное поступление)
В этой связи особенно важно, что вещества, нарушающие накопление энергии дыхания в макроэргических фосфорных связях (динитрофенол), также тормозят поступление солей. Поступление воды и солей во многих случаях идет независимо друг от друга
Сопоставление количества воды, испаренной в процессе транспирации, и количества поступивших солей показывает, что прямой зависимости между этими процессами обычно нет. Так, при выращивании растений в условиях высокой влажности воздуха транспирация резко падает, а поступление солей идет с достаточной интенсивностью.
Относительная независимость поступления воды и солей хорошо видна из следующих данных. В молодом возрасте (до фазы выхода в трубку) растения пшеницы поглощают фосфора в 14 раз больше, а в фазу колошения в два раза меньше, чем следовало бы ожидать, исходя из возможного содержания в испарившейся воде. Все же в некоторых случаях увеличение интенсивности транспирации может сказаться положительно на поглощении солей. Известно, что усиление транспирации приводит к ускорению передвижения восходящего тока воды с растворенными солями, что способствует быстрому освобождению от них клеток корня, а следовательно, косвенно ускоряет поглощение. Следует иметь в виду и трудности разграничения влияния транспирации от фотосинтеза. Открытие устьиц вызывает усиление, как того, так и другого процесса. Увеличение интенсивности фотосинтеза приводит к возрастанию содержания углеводов и, как следствие, к увеличению интенсивности дыхания и поступления солей. Ускорение темпов роста увеличивает использование питательных веществ и тем самым усиливает их поступление. Наряду с этим быстрый рост корневой системы оказывает прямое влияние на поглощение благодаря увеличению поверхности, соприкасающейся с почвой.







Корневая система и транспорт солей
13 января, 2011
На заре эволюции всем клеткам растительного организма была присуща способность к поглощению воды и питательных веществ. По-видимому, разделение функций между клетками отдельных органов осуществлялось постепенно. Это подтверждается тем, что клетки листа сохранили в определенной мере способность к поглощению не только воды, но и питательных солей. Сформировавшаяся корневаясистема — сложный специализированный орган. Основной зоной поглощения питательных веществ, снабжающей и надземные органы растения, является зона растяжения клеток и зона корневых волосков. Подсчеты показывают, что на 1 мм2 поверхности корня развивается от 200 до 400 корневых волосков. Таким образом, корневые волоски увеличивают поверхность корня в сотни раз. Они обладают и повышенной способностью к поглощению (Д.Б. Вахмистров). В меристематической зоне нет дифференцированной сосудистой системы. При этом флоэма дифференцируется раньше, и лишь несколько выше по длине корня образуется ксилема. Именно по ксилеме происходит передвижение воды с растворенными питательными веществами. Поэтому основная масса поглощенных меристемой ионов используется в этих же клетках.
Однако некоторое количество ионов, и особенно Са2+ все же поступает из этой зоны в надземные органы растений. Вместе с тем поглощенные в зоне растяжения и зоне корневых волосков ионы, наряду со снабжением надземных органов, могут транспортироваться и вниз по корню (М.Ф. Данилова). Выше зоны корневых волосков расположена зона ветвления корня. В этой зоне поверхность покрыта слоем пробки и в поглощении питательных солей практически не участвует. Различные зоны корня поглощают разные минеральные элементы. Показано, что Са2+ поступает только в апикальные зоны, К+, NH4+, фосфаты абсорбируются всей корневой системой. Однако для кукурузы установлено, что наибольшая скорость аккумуляции К+ и N03— наблюдается в зоне растяжения, a NH4+ быстрее абсорбируется в апексах.
Особенности транслокации по флоэме растения. Строение ситовидных трубок растения.
Прежде чем рассматривать возможные механизмы транслокации по флоэме, полезно перечислить некоторые факты, которые не должны противоречить любой выдвигаемой гипотезе.
1. Количество транспортируемых флоэмой растворенных веществ очень велико. Подсчитано, например, что вниз по стволу крупного дерева за вегетационный период перемешается до 250 кг сахара.
3. Транспорт может осуществляться на очень большие расстояния. Эвкалипты достигают в высоту более 100 м. Листья этих деревьев располагаются главным образом у вершины, а значит, ассимиляты должны перемещаться вниз почти по всей длине ствола, а часто еще и на значительное расстояние по корням.
4. Относительная масса флоэмы невелика. Толщина слоя функционально активной флоэмы, расположенного по окружности древесного ствола, близка к толщине почтовой открытки. Флоэма образует самый внутренний слой коры (точнее — ее луба) одревесневших стеблей и корней, при этом более старые слои флоэмы растягиваются и отмирают по мере роста органов и увеличения их диаметра.
5. Флоэмный сок движется у цветковых растений по ситовидным трубкам, диаметр которых очень мал — не более 30 мкм (как у тончайшего человеческого волоса). Через примерно равные интервалы эти трубки разделены ситовидными пластинками со сквозными отверстиями еще меньшего диаметра. Чем меньше диаметры трубок и отверстий, тем больше сопротивление потоку жидкости и тем большая сила нужна для приведения ее в движение. Давление внутри ситовидных трубок велико.
6
Помимо ситовидных пластинок, ситовидные трубки обладают другими структурными особенностями, которые также должны приниматься во внимание
Строение ситовидных трубок растения
Строение флоэмы по данным световой микроскопии описано в статье. Эта ткань содержит проводящие трубки, называемые ситовидными, которые образованы клетками — члениками ситовидных трубок, — соединенными в ряд своими торцами. Членики отделены друг от друга торцевыми ситовидными пластинками с отверстиями, позволяющими жидкости перетекать из клетки в клетку1.
В отличие от сосудов ксилемы, представляющих собой мертвые полые трубки, по которым раствор течет, почти или вообще не встречая никаких препятствий, ситовидные трубки флоэмы являются живыми, и движение растворов по ним затруднено из-за наличия ситовидных пластинок и в меньшей степени из-за наличия цитоплазмы. На рисунке приведена электронная микрофотография зрелого членика ситовидной трубки, а на рисунке — схема с указанием всех основных деталей ситовидных элементов и примыкающих к ним клеток-спутниц.
В процессе развития ситовидного элемента из меристематической клетки ядро этой клетки дегенерирует, и перед нами оказывается неооычныи пример живой клетки, не имеющей ядра; в этом отношении она сходна с эритроцитом млекопитающего. Одновременно происходит множество других важных изменений, результаты которых схематически представлены на рисунке. Клеточная стенка на обоих «торцах» членика превращается в ситовидные пластинки. Здесь плазмодесмы, соединяющие между собой соседние цитопласты, сильно утолщаются, образуя тем самым многочисленные ситовидные поры, сквозь которые они проходят. Вид ситовидной пластинки с поверхности показан на рисунке. Конечный итог всех этих преобразований — формирование трубчатой структуры, выстланной тонким пристенным слоем живой цитоплазмы, окруженной плазмалеммой. Центральная часть ситовидной трубки занята как бы единой гигантской вакуолью, которая, впрочем, не отделена от цитоплазмы тонопластом.
К каждому членику прилегают одна или несколько клеток-спутниц, которые возникают из той же самой родительской клетки путем ее продольного деления. Клетки-спутницы имеют очень плотную цитоплазму с ядром, мелкими вакуолями и обычными клеточными органелла-ми. Судя по многочисленным митохондриям и рибосомам, метаболически клетки-спутницы весьма активны. В физиологическом отношении они очень тесно связаны с ситовидными элементами и совершенно необходимы для их жизнедеятельности: в случае гибели клеток-спутниц погибают и ситовидные элементы. У некоторых растений в ситовидных элементах образуется большое количество волокнистого белка, называемого флоэмным белком (Ф-белком). Иногда он образует крупные отложения, различимые в световом микроскопе. Раньше его функция вызывала много споров, но сейчас признано, что особой роли в транслокации он не играет.
Механизм флоэмного транспорта
7 декабря, 2010
В 1930 г. Э. Мюнх выдвинул гипотезу «массового тока», согласно которой по ситовидным трубкам из одного членика в другой через поры движется жидкость с растворенными веществами. Движущей силой этого потока является тургорное давление. Клетки, в которых образуются сахара (вещество-донор), характеризуются высокой концентрацией клеточного сока и высоким тургорным давлением, а клетки, в которых сахара потребляются,— низким тургорным давлением (структура-акцептор). Если эти клетки соединены между собой, то жидкость должна перетекать из клеток с высоким давлением в клетки с низким давлением. Гипотеза Мюнха в последнее время приобрела сторонников, однако она не согласуется с многими факторами. Далеко не всегда передвижение идет по градиенту тургорного давления (в сторону его уменьшения). Эта гипотеза не объясняет как происходит переброска ассимилятов из опадающих листьев или завядающих лепестков цветка, которые обладают, низким тургорным давлением. Расчеты показывают также, что для передвижения раствора сахарозы с той скоростью, которая наблюдается в ситовидных трубках, нужна сила, значительно превосходящая силу тургорного давления, развиваемую в клетках-донорах. Альтернативной является гипотеза, согласно которой передвижение органических веществ идет с затратой энергии. По данным А.Л. Курсанова, существует взаимосвязь между флоэмным транспортом и напряженностью энергетического обмена. Источником энергии для транспорта веществ может быть АТФ, образовавшаяся как в самих ситовидных элементах, так и, главным образом, в клетках-спутницах. Известно, что клетки-спутницы характеризуются исключительно высокой интенсивностью дыхания и окислительного фосфорилирования. Возможно, что периодические сокращения белковых тяжей ситовидных трубок могут способствовать перемещению веществ в определенном направлении. Ряд исследователей говорят о электроосмотических явлениях при транспорте веществ по флоэме. Согласно этой гипотезе, на каждой ситовидной пластинке возникает электрический потенциал. Возникновение электрического потенциала связано с циркуляцией ионов К+. Предполагается, что К+ активно (с затратой энергии АТФ) поглощается выше ситовидной перегородки и проникает через нее в нижний членик. По другую сторону перегородки ионы К+ пассивно выходят в сопровождающую клетку. Активное поступление К+ с одной стороны ситовидной трубки обеспечивается тем, что ассимиляционный поток обогащает ситовидную трубку АТФ именно с той стороны, к которой он первоначально подходит. Возникающий на каждой ситовидной пластинке электрический потенциал и является движущей силой потока сахарозы по флоэме




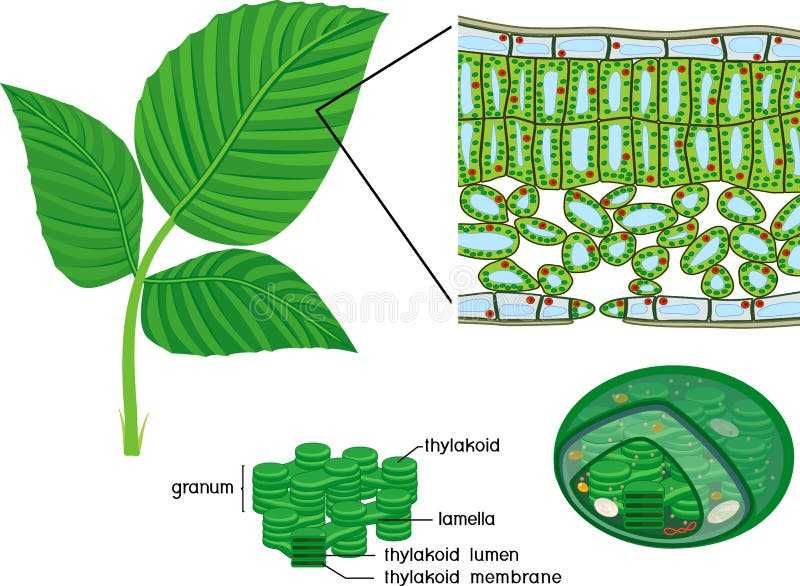
![Направление движения питательных веществ по флоэме [1983 гэлстон а., дэвис п., сэттер р. - жизнь зеленого растения]](https://lesniepolyani.ru/wp-content/uploads/f/c/7/fc76b686ff2cac8b5de6d44227339fe2.jpeg)


Важно отметить, что флоэма, имеет и вторую функцию — распространение импульсов возбуждения, что может являться одним из механизмов, обеспечивающих транспорт (В.А. Опритов)
Д. Фенсом выдвинута гипотеза, что транспорт ассимилятов по флоэме осуществляется с помощью нескольких механизмов. Основное значение при этом придается тем механизмам, которые были рассмотрены, т.е. перетеканию раствора под давлением, электроосмотической гипотезе и передвижению, связанному с перистальтическим сокращением белковых тяжей. Каждый из этих механизмов может осуществляться и усиливаться при отсутствии условий для функционирования других. Важное значение имеет направление движения и распределение ассимилятов. Оно во многом определяется интенсивностью использования веществ, потребностями того или иного органа, интенсивностью его роста.
Механизм почвенного питания
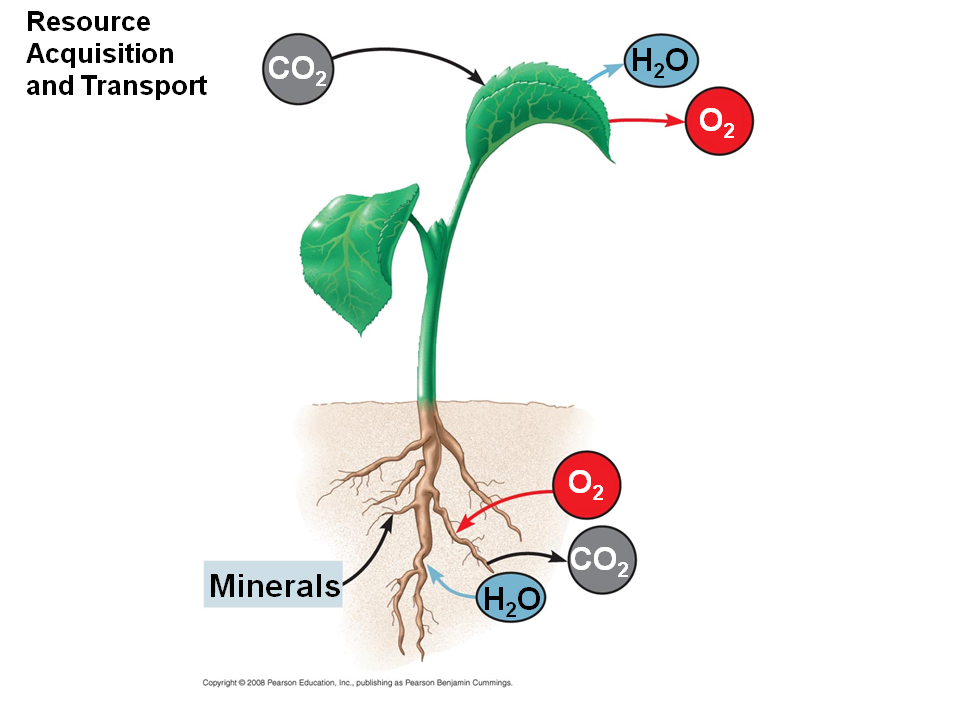
Таким образом, в растении одновременно осуществляется передвижение веществ в противоположных направлениях. В ботанике этот процесс называют восходящим и нисходящим током.
Но какие силы заставляют воду из почвы двигаться вверх? Оказывается, что это происходит под влиянием корневого давления и транспирации — испарения воды с поверхности листьев.
Для растений этот процесс является жизненно необходимым. Дело в том, что только в почве находятся минералы, без которых развитие тканей и органов будет невозможным. Так, азот необходим для развития корневой системы. В воздухе этого элемента предостаточно — 75 %
Но растения не способны фиксировать атмосферный азот, поэтому минеральное питание так важно для них
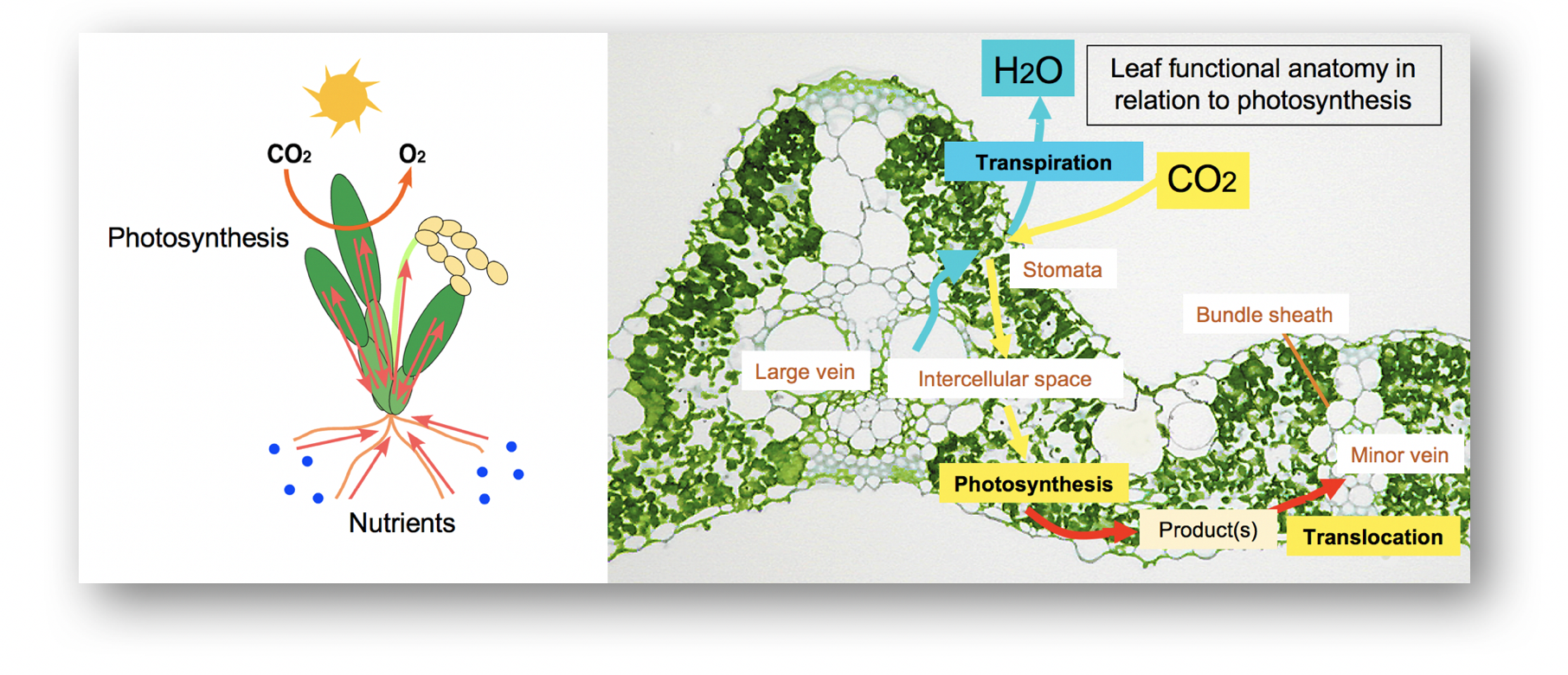
Поднимаясь, молекулы воды плотно сцепляются между собой и стенками сосудов. При этом возникают силы, способные поднять воду на приличную высоту — до 140 м. Такое давление заставляет почвенные растворы через корневые волоски проникать в кору, и далее к сосудам ксилемы. По ним вода поднимается к стеблю. Далее, под действием транспирации, вода поступает в листья.
В жилках рядом с сосудами находятся и ситовидные трубки. Эти элементы осуществляют нисходящий ток. Под воздействием солнечного света в хлоропластах листа синтезируется полисахарид глюкоза. Это органическое вещество растение расходует на осуществление роста и процессов жизнедеятельности.
Итак, проводящая ткань растения обеспечивает передвижение водных растворов органических и минеральных веществ по растению. Ее структурными элементами являются сосуды и ситовидные трубки.
Загрузка ситовидных трубок растения.
Было показано, что концентрация сахарозы в ситовидных трубках листьев обычно составляет от 10 до 30%, тогда как в фотосинтезирующих клетках она держится на уровне 0,5%. Следовательно, как мы уже отмечали, загрузка флоэмы идет против осмотического градиента. Этот механизм активно изучался в последние годы. Сначала растворенные органические вещества должны переместиться из хлоропластов во флоэму; этот путь составляет не более 3 мм, причем вещества могут двигаться и по симпласту, и по апопласту.
Симпластный путь предполагает перемещение веществ по плазмодесмам, а апопластный — через клеточные стенки посредством диффузии или объемного потока в транспирационном паре.
В 1968 г. Гуннинг и его сотрудники описали новый тип модифицированных клеток-спутниц — передаточные клетки. Как видно из рисунка, эти клетки примыкают к ситовидным трубкам. В результате дополнительного неравномерного утолщения клеточные стенки у них образуют многочисленные внутренние выступы, что почти в десять раз увеличивает площадь выстилающей такую стенку плазмалеммы. Как полагают, такое видоизменение клеток связано с необходимостью активно поглощать растворенные вещества из соседних клеток. Энергию для всех активных процессов дают многочисленные митохондрии передаточных клеток. Передаточные клетки обнаружены не у всех растений, но они обычны у бобовых и некоторых других семейств. Впрочем, считают, что активный транспорт происходит и в их отсутствие.
Как полагают, загрузку сахарозы (и ряда других компонентов раствора, в том числе аминокислот, фосфатов, ионов калия и аммония) в клетки-спутницы осуществляют особые белки-переносчики, локализованные в плазмалемме этих клеток. Образуемые белками системы переноса сходны с аналогичными системами животных и бактериальных клеток, где транспорт органических молекул сопряжен с транспортом протонов (Н+). Протоны выкачиваются из клетки переносчиком, использующим для этого энергию АТФ. В результате формируется градиент концентрации Н+, генерирующий потенциальную энергию. Протоны быстро диффундируют назад в клетку с участием других белков-переносчиков, которые одновременно пропускают внутрь молекулы сахарозы или других органических веществ (котранспорт по типу сим-порта). Таким образом, движущая сила этого процесса — формирование протонного градиента через плазмалемму с пониженным рН (более высокой концентрацией Н+) вне клетки.
В результате активного транспорта веществ в клетки-спутницы осмотический потенциал в них сильно понижается. Это стимулирует поступление в них воды путем осмоса, повышение давления и движение растворов (в том числе и раствора сахарозы) по механизму объемного потока через многочисленные плазмодесмы, связывающие клетки-спутницы с ситовидными трубками. Таким образом высокое давление и объемный поток создаются в ситовидных трубках, а не в клетках мезофилла листа, как предполагал Мюнх. Возможно также, что механизм активного транспорта через плазмодесмы из клеток-спутниц в ситовидные трубки точно такой же, как и механизм, обеспечивающий поступление сахарозы в клетки-спутницы.
При разгрузке ситовидных трубок растворенные вещества, вероятно, уходят из них в передаточные клетки по плазмодесмам. В результате водный потенциал трубок повышается (становится менее отрицательным). Это тормозит потенциальный осмос извне и поддерживает гидростатический градиент между местами загрузки и разгрузки флоэмы. Полагают, что последний процесс может идти и через плазмалем-му ситовидных трубок в их клеточные стенки и далее апопластным путем.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним. См. подробнее в пользовательском соглашении.
Ситовидные трубки растений
Это проводящие клетки луба. Между собой они разделены многочисленными перегородками. Внешне их строение напоминает сито. Отсюда и происходит название. Ситовидные трубки растений живые. Это объясняется слабым давлением нисходящего тока.


Их поперечные стенки пронизаны густой сетью отверстий. А клетки содержат много сквозных отверстий. Все они являются прокариотическими. Это означает, что в них нет оформленного ядра.
Живыми элементы цитоплазмы ситовидных трубок остаются только на определенное время. Продолжительность этого периода варьирует в широких пределах — от 2 до 15 лет. Данный показатель зависит от вида растения и условий его произрастания. Ситовидные трубки транспортируют воду и органические вещества, синтезированные в процессе фотосинтеза от листьев к корню.
Влияние внешних условий на транспорт веществ растений
13 января, 2011
При температуре, близкой к 0°С, поглощение солей идет медленно, затем, в пределах до 40°С, оно усиливается. Увеличение температуры на 10°С может вызвать возрастание поглощения в два и даже в три раза. В темноте поглощение солей замедляется и постепенно прекращается, а под влиянием освещения ускоряется. Так, при освещении поглощение фосфора усиливается уже через 2—5 мин. Быстрота реакции указывает на прямое действие света. Вместе с тем свет может оказывать и косвенное влияние. На свету в процессе фотосинтеза образуются углеводы, которые необходимы для дыхания. Зависимость поглощения от интенсивности дыхания проявляется четко. При длительном выдерживании растений в темноте, после того как запас дыхательных субстратов исчерпан, поглощение солей не только прекращается, но может даже наблюдаться их выделение. Нельзя также не учитывать, что на свету в процессе фотофосфорилирования образуется АТФ, энергия которой используется на поступление веществ. При уменьшении содержания кислорода до 2—3% интенсивность поступления солей остается на одном уровне. Лишь снижение концентрации кислорода ниже 3% вызывает падение поглощения примерно в два раза. Необходимо учитывать, что и интенсивность дыхания сохраняется на высоком уровне в широком диапазоне концентраций кислорода. Падение интенсивности дыхания наблюдается при той же концентрации кислорода, при которой поглощение солей уменьшается. Концентрация ионов водорода (рН) также сказывается на поглощении солей.
Еще в опытах Д.А. Сабинина и И.И. Колосова было показано, что при подкислении раствора поступление катионов задерживается. Это происходит, по всей вероятности, в силу конкуренции, которая наблюдается между одинаково заряженными ионами за возможность вступить в реакцию с переносчиками. Вместе с тем подкисление улучшает доступность ионов фосфорной кислоты. Наоборот, подщелачивание внешнего раствора снижает поступление фосфора благодаря переходу одновалентного иона Н2Р04— в двухвалентный НР04 2- и трехвалентный Р043-, которые являются менее доступными для растения. Резкоеизменение значения рН может также оказать влияние из-за повреждения мембран клетки. Как уже упоминалось, поглощение одного иона зависит от присутствия других ионов. Так, в присутствии легко поглощаемого аниона катионы той же соли поступают быстрее. Ионы с одинаковым зарядом обычно конкурируют между собой. Однако в некоторых случаях наблюдается противоположная закономерность. Так, при наличии в среде иона фосфора (Р043-) поглощение нитратов (N03—) ускоряется. Это может явиться косвенным результатом увеличения использования нитратов в присутствии фосфорной кислоты.
ЗАКЛЮЧЕНИЕ
Результаты наших исследований показали, что величина оттока ассимилятов из листьев
картофеля определяется собственным возрастом листа, его положением на стебле и этапом
онтогенеза растения. Включение в донорно-акцепторную систему материнского клубня позволяет
существенно уменьшить потребление продуктов текущего фотосинтеза на формирование и
поддержание функциональной активности листьев нижнего яруса и тем самым обеспечить
создание временного фонда ассимилятов, используемых при клубнеобразовании. Листья
среднего яруса потребляют на собственный рост и обновление структур 30–35% ассимилированного
ими углерода, тогда как у нижних листьев эта величина втрое меньше. Инициация клубнеобразования
канализирует транспорт и запасание ассимилятов. В клубнях локализуется как минимум
50–55% первоначально ассимилированного растением углерода. Воздействие коротким днем
стимулировало распределение ассимилированного углерода в клубни у растений картофеля
с количественно-короткодневной реакцией клубнеобразования, тогда как у сортового картофеля
с нейтральной реакцией сокращение суточной продолжительности фотосинтеза приводило
к торможению накопления биомассы, но не оказывало существенного влияния на характер
распределения ассимилятов. В целом, полученные данные дополняют представления об организации
и функционировании системы ДАС и обеспечивают основу для разработки способов управления
ДАО картофеля в целях повышения продуктивности этой важнейшей сельскохозяйственной
культуры.
Финансовое обеспечение работы осуществлялось из средств федерального бюджета на выполнение
государственного задания № АААА-А17-117033010038-7 и АААА-А18-118012290132-0.