

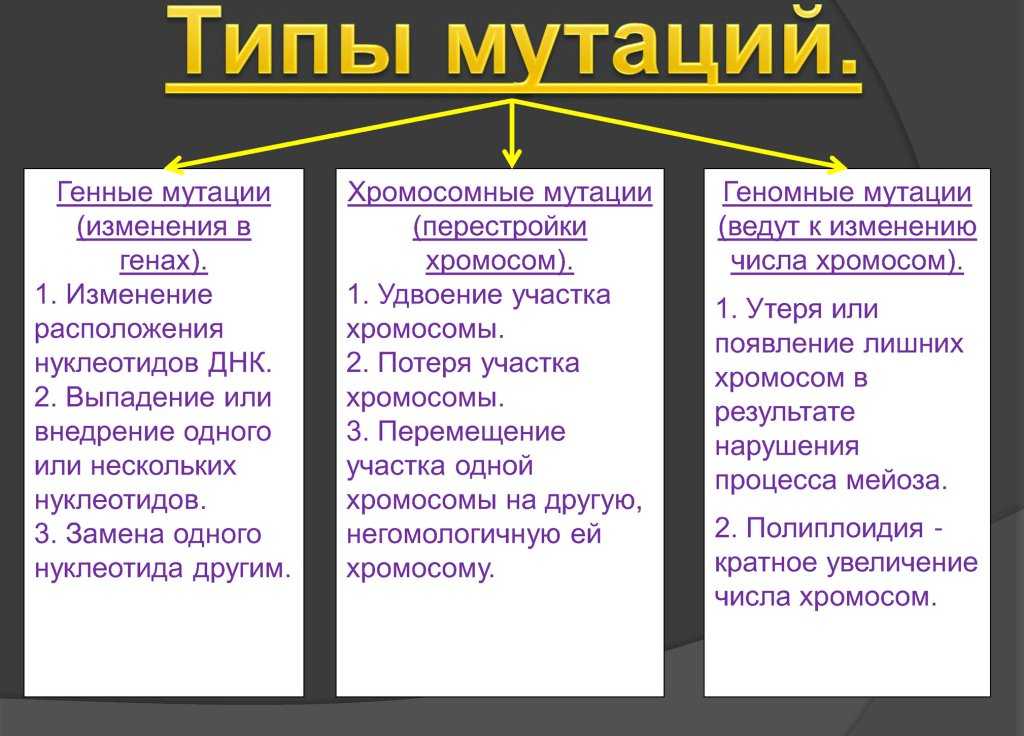
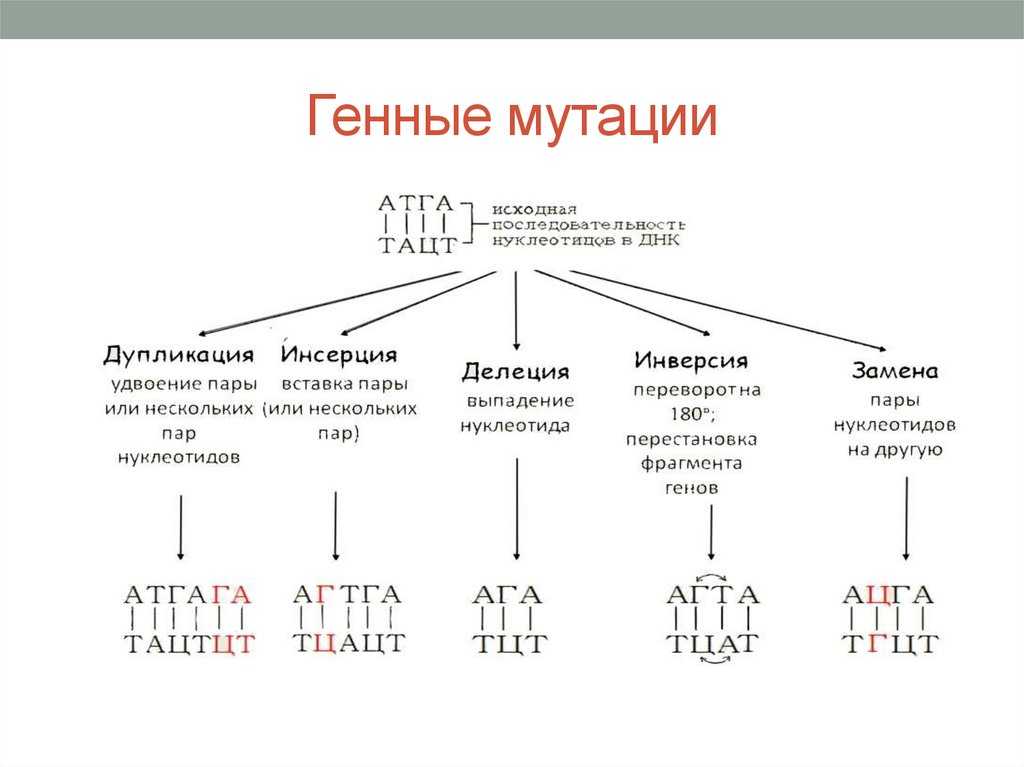
Делеции нуклеотидов. Инсерции нуклеотидов. Мутации сдвига рамки считывания. Кроссинговер. Неравный кроссинговер.
Если однонуклеотидные замены, делеции или инсерции нуклеотидов происходят в промоторной области гена, то такие мутации способны нарушать взаимодействие РНК-полимеразы и других белков, участвующих в инициации транскрипции, что приведет к уменьшению или даже прекращению синтеза мРНК на матрице ДНК.
Рис. 4.1. Механизм возникновения дупликации или делеции гена РМР22. а — нормальное спаривание гомологичных участков хромосомы 17р, в которых расположен ген РМР22. По краям участка расположены гомологичные повторы длиной около 30 тыс. п.н.; б — негомологичное спаривание участков хромосомы 17р за счет гомологичных повторов и неравный кроссинго-вер в спаренных гомологичных участках; в — в результате неравного крос-синговера происходят удвоение гена РМР22 в одном из гомологов и его де-леция во втором гомологе (на рис. 4.1 делеция не показана).
Следующий класс молекулярных мутаций — это делеции (утраты) или инсерции (вставки) нуклеотидов. В том случае, когда делетируется или вставляется тройка нуклеотидов, то если этот триплет является кодирующим, в составе полипептида либо исчезает определенная аминокислота, либо появляется новая аминокислота. Однако, если в результате делеции или инсерции вставляется или удаляется число нуклеотидов, не кратное трем, то меняется или утрачивается смысл для всех остальных, следующих за вставкой или делецией кодонов молекулы мРНК. Такие мутации называются мутациями сдвига рамки считывания. Нередко они приводят к образованию стоп-кодона в следующей за инсерцией или делецией последовательности нуклеотидов мРНК.
На схеме показано, что мутация сдвига рамки считывания возникла в результате инсерции пары оснований. Это привело к изменению кодонов после инсерции и аминокислотной последовательности полипептидной цепи.
Одним из механизмов возникновения мутаций является неравный кроссинговере. По-видимому, именно таким образом возникает, например, большая часть делеций/дупликаций в области локализации гена РМР22 на хромосоме, кодирующего периферический белок миелина 22. Участок хромосомы 17, в котором расположен ген РМР22 протяженностью примерно в 1,5 млн п.н., фланкируется двумя высокогомологичными повторами, каждый длиной около 30 тыс. п.н.
P.S. Кроссинговером называется обмен генетическим материалом между гомологичными хромосомами в мейозе I.
Эволюционные последствия
При неравном кроссинговере между хромосомами может происходить большой обмен последовательностями. По сравнению с генной конверсией, которая может передавать максимум 1500 пар оснований, было обнаружено, что неравный кроссинговер в генах дрожжевой рДНК переносит около 20000 пар оснований в одном событии кроссовера. Неравный кроссовер может сопровождаться согласованной эволюцией дублированных последовательностей.
Было высказано предположение, что более длинный интрон, обнаруженный между двумя генами бета-глобина, является ответом на вредный отбор в результате неравного кроссинговера в генах бета-глобина. Сравнение альфа-глобина, не имеющего длинных интронов, и генов бета-глобина показывает, что согласованная эволюция альфа-глобина в 50 раз выше.
Когда неравный кроссинговер создает дупликацию гена , у дубликата есть 4 эволюционных судьбы. Это связано с тем, что очищающий отбор, действующий на дублированную копию, не очень силен. Теперь, когда есть дублирующая копия, нейтральные мутации могут воздействовать на дубликат. Чаще всего нейтральные мутации будут продолжаться до тех пор, пока дубликат не станет псевдогеном . Если дубликат копии увеличивает эффект дозировки продукта гена, то дубликат может быть сохранен как дублирующая копия. Неофункционализация также возможна: дублированная копия приобретает мутацию, которая придает ей другую функцию, чем ее предок. Если обе копии приобретают мутации, возможно, что происходит субфункциональное событие . Это происходит, когда обе дублированные последовательности выполняют более специализированную функцию, чем предковая копия.
Размер генома
Дупликации генов являются основной причиной увеличения размера генома, и, поскольку неравный кроссинговер является основным механизмом дупликации генов, неравный кроссинговер способствует эволюции размера генома, что является наиболее распространенным событием региональной дупликации, которое увеличивает размер генома.
Мусорная ДНК
При просмотре генома эукариота бросается в глаза большое количество тандемных повторяющихся последовательностей ДНК, составляющих большую часть генома. Например, более 50% генома Dipodmys ordii состоит из трех специфических повторов. Drosophila virilis имеет три последовательности, которые составляют 40% генома, а 35% Absidia glauca представляют собой повторяющиеся последовательности ДНК. На эти короткие последовательности не действует давление отбора, и частота повторов может быть изменена неравным кроссинговером.
Причины мутаций[]
Мутации делятся на спонтанные и индуцированные. Спонтанные мутации возникают самопроизвольно на протяжении всей жизни организма в нормальных для него условиях окружающей среды с частотой около 10 в −9 степени — 10 в −12 на нуклеотид за клеточную генерацию.
Индуцированными мутациями называют наследуемые изменения генома, возникающие в результате тех или иных мутагенных воздействий в искусственных (экспериментальных) условиях или при неблагоприятных воздействиях окружающей среды.
Мутации появляются постоянно в ходе процессов, происходящих в живой клетке. Основные процессы, приводящие к возникновению мутаций — репликация ДНК, нарушения репарации ДНК и генетическая рекомбинация.
Связь мутаций с репликацией ДНК
Многие спонтанные химические изменения нуклеотидов приводят к мутациям, которые возникают при репликации. Например, из-за деаминирования цитозина напротив него в цепь ДНК может включаться урацил (образуется пара У-Г вместо канонической пары Ц-Г). При репликации ДНК напротив урацила в новую цепь включается аденин, образуется пара У-А, а при следующей репликации она заменяется на пару Т-А, то есть происходит транзиция.
Генетические карты хромосом
Большая советская энциклопедия. — М.: Советская энциклопедия . 1969—1978 .
Смотреть что такое «Генетические карты хромосом» в других словарях:
ГЕНЕТИЧЕСКИЕ КАРТЫ ХРОМОСОМ — схемы относительного расположения генов в хромосомах, позволяющие предсказывать характер наследования изучаемых признаков организмов … Большой Энциклопедический словарь
генетические карты хромосом — схемы относительного расположения генов в хромосомах, позволяющие предсказывать характер наследования изучаемых признаков организмов. * * * ГЕНЕТИЧЕСКИЕ КАРТЫ ХРОМОСОМ ГЕНЕТИЧЕСКИЕ КАРТЫ ХРОМОСОМ, схемы относительного расположения генов в… … Энциклопедический словарь
ГЕНЕТИЧЕСКИЕ КАРТЫ ХРОМОСОМ — схемы относительного расположения генов в хромосомах, позволяющие предсказывать характер наследования изучаемых признаков организмов … Естествознание. Энциклопедический словарь
Карты хромосом — Графическое представление нормального человеческого кариотипа. Генетические карты хромосом это схема взаимного расположения и относительных расстояний между генами определенных хромосом, находящихся в од … Википедия
Цитологические карты хромосом — схематическое изображение хромосом (См. Хромосомы) с указанием мест фактического размещения отдельных генов, полученное с помощью цитологических методов. Ц. к. х. составляют для организмов, для которых обычно уже имеются Генетические… … Большая советская энциклопедия






Хромосомные карты генетические к — Хромосомные карты, генетические к. * храмасомныя карты, генетычныя к. * chromosome maps or genetic m. графическое изображение хромосом, как правило, в виде прямых линий, на которых указаны гены/локусы определенной группы сцепления (см.) и… … Генетика. Энциклопедический словарь
Генетика — I Генетика (от греч. génesis происхождение) наука о законах наследственности и изменчивости организмов. Важнейшая задача Г. разработка методов управления Наследственностью и наследственной Изменчивостью для получения нужных человеку форм… … Большая советская энциклопедия
Генетика — I Генетика (от греч. génesis происхождение) наука о законах наследственности и изменчивости организмов. Важнейшая задача Г. разработка методов управления Наследственностью и наследственной Изменчивостью для получения нужных человеку форм… … Большая советская энциклопедия
генетика — , и; ж. . Наука о законах наследственности и изменчивости организмов. Г. человека. Г. растений. Медицинская г. Космическая г. * * * генетика (от греч. génesis происхождение), наука о… … Энциклопедический словарь
Хромосомная теория наследственности — Хромосомная теория наследственности теория, согласно которой хромосомы, заключённые в ядре клетки, являются носителями генов и представляют собой материальную основу наследственности, то есть преемственность свойств организмов в ряду… … Википедия
Инсерции. Что такое инсерция? Инверсии. Что такое инверсия?
Когда сегмент одной хромосомы переносится и вставляется в другую хромосому, такую перестройку называют инсерцией.
Для того чтобы произошла инсерция, необходимо не менее 3 разрывов хромосом. Поскольку в случае возникновения инсерции не теряется и не добавляется новый генетический материал, такую перестройку считают сбалансированной. Однако у носителей такой инсерции 50 % гамет окажутся несбалансированными, поскольку они будут нести хромосому либо с делецией, либо с инсерцией.
Вследствие этого будут образовываться зиготы с частичной моносомией или частичной трисомией.
Перицентрическая и парацентрическая инверсии в хромосоме 6.
Инверсии. Что такое инверсия?
Инверсией называют хромосомную мутацию, когда после двух разрывов в одной хромосоме сегмент хромосомы, расположенный между разрывами, поворачивается на 180° и занимает инвертированное положение. Если в инвертированный сегмент попадает центромера, то такую инверсию называют перицентрическои, а если инверсия сегмента хромосомы происходит в пределах одного плеча — парацентрической.
При инверсии не происходит потери генетического материала, кроме тех случаев, когда разрыв хромосомы может затронуть функционально важный ген. Поэтому носители обоих типов инверсий не имеют, как правило, каких-либо патологических симптомов. Более того, некоторые инверсии, например перицентрическая инверсия в хромосоме 9, встречаются как нормальный признак с достаточно высокой частотой в некоторых этнических группах. Как и при других сбалансированных перестройках, инверсии в мейозе могут приводить к образованию несбалансированных гамет.
Это происходит тогда, когда в пределах инверсии возникает хиазма и осуществляется кроссинговер. В случае перицентрической инверсии это приводит к образованию двух комплементарных несбалансированных хромосом.
Одна из них будет иметь дупликацию дистального неинвертированного сегмента и делецию противоположного конца хромосомы, вторая — делецию дистального неинвертированного сегмента и дупликацию противоположного конца хромосомы. Кроссинговер в инвертированном сегменте при парацентрической инверсии, как видно из рисунка, приведет к образованию либо ацентрического фрагмента хромосомы, либо дицентрика.
Механизм образования рекомбинантных несбалансированных хромосом в случае перицентрической (I) и парацентрической (II) инверсии, когда кроссинговер происходит в инверсионной петле
Все 4 типа несбалансированных гамет в случае их участия в оплодотворении дадут нежизнеспособные зиготы, которые элиминируются на ранних стадиях развития. Вероятно, однако, что кроссинговер внутри инверсий может происходить крайне редко, так как конъюгация гомологичных хромосом при наличии в одной из них инверсии затруднена.
У экспериментальных животных инверсии используют как раз для запирания кроссинговера.
Проблема случайности мутаций[]
В 40-е годы среди микробиологов была популярна точка зрения, согласно которой мутации вызываются воздействием фактора среды (например. антибиотика), к которому они позволяют адаптироваться. Для проверки этой гипотезы был разработан флуктуационный тест и метод реплик. Флуктуационный тест Лурия-Дельбрюка заключается в том, что небольшие порции исходной культуры бактерий рассеивают в пробирки с жидкой средой, а после нескольких циклов делений добавляют в пробирки антибиотик. Затем (без последующих делений) на чашки Петри с твердой средой высевают выживших устойчивых к антибиотику бактерий. Тест показал. что число устойчивых колоний из разных пробирок очень изменчиво — в большинстве случаев оно небольшое (или нулевое), а в некоторых случаях очень высокое. Это означает, что мутации, вызвавшие устойчивость к антибиотику, возникали в случайные моменты времени как до, так и после его воздействия. Метод реплик (в микробиологии) заключается в том, что с исходной чашки Петри, где на твердой среде растут колонии бактерий, делается отпечаток на ворсистую ткань, а затем с ткани бактерии переносятся на несколько других чашек, где рисунок их расположения оказывается тем же, что на исходной чашке. После воздействия антибиотиком на всех чашках выживают колонии, расположенные в одних и тех же точках. Высевая такие колонии на новые чашки, можно показать, что все бактерии внутри колонии обладают устойчивостью. Таким образом, обоими методами было доказано, что «адаптивные» мутации возникают независимо от воздействия того фактора, к которому они позволяют приспособиться, и в этом смысле мутации случайны. Однако несомненно, что возможность тех или иных мутаций зависит от генотипа и канализована предшествующим ходом эволюции (см. Закон гомологических рядов в наследственной изменчивости). Кроме того, закономерно различается частота мутирования разных генов и разных участков внутри одного гена. Также известно, что высшие организмы используют «целенаправленные» (то есть происходящие в определенных участках ДНК) мутации в механизмах иммунитета. С их помощью создаётся разнообразие клонов лимфоцитов, среди которых в результате всегда находятся клетки, способные дать иммунный ответ на новую, неизвестную для организма болезнь. Подходящие лимфоциты подвергаются положительной селекции, в результате возникает иммунологическая память.
Замена азотистых оснований
Если происходит изменение только одного нуклеотида в ДНК, то такая мутация называется точечной. В случае мутаций по типу замены азотистых оснований одна комплементарная нуклеотидная пара молекулы ДНК заменяется в ряду циклов репликации на другую. Частота подобных происшествий составляет около 20% от общей массы всех генных мутаций.
Примером подобного является дезаминирование цитозина, в результате чего образуется урацил.
В ДНК образуется нуклеотидная пара Г-У, вместо Г-Ц. Если ошибка не будет репарирована ферментом ДНК-гликолазой, то при репликации произойдет следующее. Цепи разойдутся, напротив гуанина будет установлен цитозин, а напротив урацила — аденин. Таким образом, одна из дочерних молекул ДНК будет содержать аномальную пару У-А. При ее последующей репликации в одной из молекул напротив аденина будет установлен тимин. Т. е. в гене произойдет замена пары Г-Ц на А-Т.
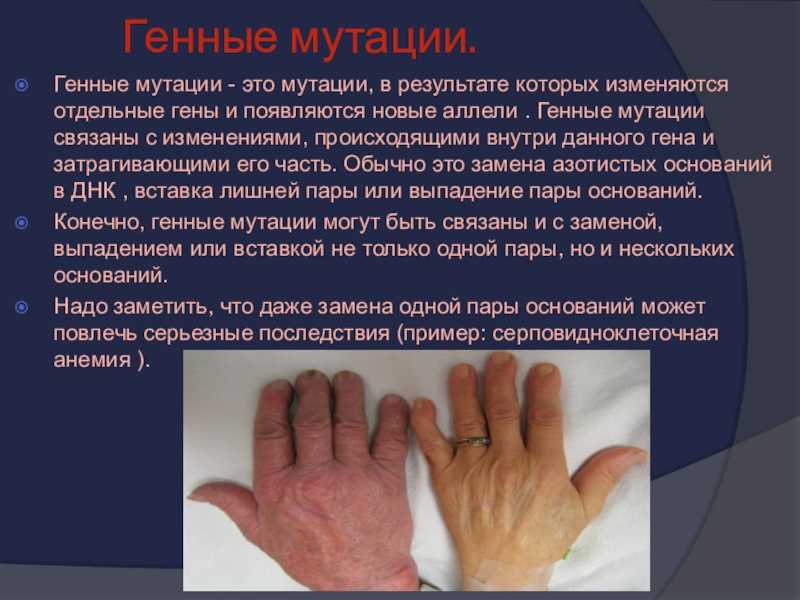






Другим примером является дезаминирование метилированного цитозина, в результате которого образуется тимин. В последствии может возникнуть ген с парой Т-А вместо Ц-Г.
Могут быть и обратные замены: пара А-Т при определенных химических реакциях может заменяться на Ц-Г. Например, в процессе репликации к аденину может присоединиться бромурацил, который при следующей репликации присоединяет к себе гуанин. В следующем цикле гуанин свяжется с цитозином. Таким образом в гене пара А-Т заменится на Ц-Г.
Замена одного пиримидина на другой пиримидин или одного пурина на другой пурин называется транзицией. Пиримидинами являются цитозин, тимин, урацил. Пуринами — аденин и гуанин. Замена пурина на пиримидин или пиримидина на пурин называется трансверсией.
Точечная мутация может не привести ни к каким последствиям из-за вырожденности генетического кода, когда несколько кодонов-триплетов кодируют одну и ту же аминокислоту. Т. е. в результате замены одного нуклеотида может образоваться другой кодон, но кодирующий ту же аминокислоту, что и старый. Такая замена нуклеотидов называется синонимической. Их частота около 25% от всех замен нуклеотидов. Если же смысл кодона меняется, он начинает кодировать другую аминокислоту, то замена называется мисенс-мутацией. Их частота около 70%.
В случае мисенс-мутации при трансляции в пептид будет включена не та аминокислота, в результате чего его свойства изменятся. От степени изменения свойств белка зависит степень изменения более сложных признаков организма. Например, при серповидно-клеточной анемии в белке заменена лишь одна аминокислота — глутамин на валин. Если же глутамин заменяется на лизин, то свойства белка меняются не сильно, т. е. обе аминокислоты гидрофильны.
Точечная мутация может быть такой, что на месте кодирующего аминокислоту кодона возникает стоп-кодон (УАГ, УАА, УГА), прерывающий (терминирующий) трансляцию. Это нонсенс-мутации. Иногда бывают и обратные замены, когда на месте стоп-кодона возникает смысловой. При любой подобной генной мутации функциональный белок уже не может быть синтезирован.
Роль мутаций в эволюции[]
При существенном изменении условий существования те мутации, которые раньше были вредными, могут оказаться полезными. Таким образом, мутации являются материалом для естественного отбора. Так, мутанты-меланисты (темноокрашенные особи) в популяциях березовой пяденицы (Biston betularia) в Англии впервые были обнаружены учеными среди типичных светлых особей в середине XIX века. Темная окраска возникает в результате мутации одного гена. Бабочки проводят день на стволах и ветвях деревьев, обычно покрытых лишайниками, на фоне которых светлая окраска является маскирующей. В результате промышленной революции, сопровождающейся загрязнением атмосферы, лишайники погибли, а светлые стволы берез покрылись копотью. В результате к середине XX века (за 50-100 поколений) в промышленных районах темная морфа почти полностью вытеснила светлую. Было показано, что главная причина преимущественного выживания черной формы — хищничество птиц, которые избирательно выедали светлых бабочек в загрязненных районах.
Если мутация затрагивает «молчащие» участки ДНК, либо приводит к замене одного элемента генетического кода на синонимичный, то она обычно никак не проявляется в фенотипе (проявление такой синонимичной замены может быть связано с разной частотой употребления кодонов). Однако методами генного анализа такие мутации можно обнаружить. Поскольку чаще всего мутации происходят в результате естественных причин, то в предположении, что основные свойства внешней среды не менялись, получается, что частота мутаций должна быть примерно постоянной. Этот факт можно использовать для исследования филогении — изучения происхождения и родственных связей различных таксонов, в том числе и человека. Таким образом, мутации в молчащих генах служат для исследователей своеобразными «молекулярными часами». Теория «молекулярных часов» исходит также из того, что большинство мутаций нейтральны, и скорость их накопления в данном гене не зависит или слабо зависит от действия естественного отбора и потому остается постоянной в течение длительного времени. Для разных генов эта скорость, тем не менее, будет различаться.
Исследование мутаций в митохондриальной ДНК (наследуется по материнской линии) и в Y-хромосомах (наследуется по отцовской линии) широко используется в эволюционной биологии для изучения происхождения рас и народностей, реконструкции биологического развития человечества.
Последствия мутаций для клетки и организма[]
Мутации, которые ухудшают деятельность клетки в многоклеточном организме, часто приводят к уничтожению клетки (в частности, к программируемой смерти клетки, — апоптозу). Если внутри- и внеклеточные защитные механизмы не распознали мутацию и клетка прошла деление, то мутантный ген передастся всем потомкам клетки и, чаще всего, приводит к тому, что все эти клетки начинают функционировать иначе.
Мутация в соматической клетке сложного многоклеточного организма может привести к злокачественным или доброкачественным новообразованиям, мутация в половой клетке — к изменению свойств всего организма-потомка.
В стабильных (неизменных или слабо изменяющихся) условиях существования большинство особей имеют близкий к оптимальному генотип, а мутации вызывают нарушение функций организма, снижают его приспособленность и могут привести к смерти особи. Однако в очень редких случаях мутация может привести к появлению у организма новых полезных признаков, и тогда последствия мутации оказываются положительными; в этом случае они являются средством адаптации организма к окружающей среде и, соответственно, называются адаптационными.
Классификации мутаций[]
Существует несколько классификаций мутаций по различным критериям. Мёллер предложил делить мутации по характеру изменения функционирования гена на гипоморфные (измененные аллели действуют в том же направлени, что и аллели дикого типа; синтезируется лишь меньше белкового продукта), аморфные (мутация выглядит, как полная потеря функции гена, например, мутация white у Drosophila), антиморфные (мутантный признак изменяется, например, окраска зерна кукурузы меняется с пурпурного на бурый) и неоморфные.
В современной учебной литературе используется и более формальная классификация, основанная на характере изменения структуры отдельных генов, хромосом и генома в целом. В рамках этой классификации различают следующие виды мутаций:
- генные
- хромосомные
- геномные.
Геномные: — полиплоидизация (образование организмов или клеток, геном которых представлен более чем двумя (3n, 4n, 6n и т. д.) наборами хромосомами) и анеуплоидия (гетероплоидия) — изменение числа хромосом, не кратное гаплоидному набору (см. Инге-Вечтомов, 1989). В зависимости от происхождения хромосомных наборов среди полиплоидов различают аллополиплоидов, у которых имеются наборы хромосом, полученные при гибридизации от разных видов, и аутополиплоидов, у которых происходит увеличение числа наборов хромосом собственного генома, кратное n.
При хромосомных мутациях происходят крупные перестройки структуры отдельных хромосом. В этом случае наблюдаются потеря (делеция) или удвоение части (дупликация) генетического материала одной или нескольких хромосом, изменение ориентации сегментов хромосом в отдельных хромосомах (инверсия), а также перенос части генетического материала с одной хромосомы на другую (транслокация) (крайний случай — объединение целых хромосом, т.н. Робертсоновская транслокация, которая является переходным вариантом от хромосомной мутации к геномной).
На генном уровне изменения первичной структуры ДНК генов под действием мутаций менее значительны, чем при хромосомных мутациях, однако генные мутации встречаются более часто. В результате генных мутаций происходят замены, делеции и вставки одного или нескольких нуклеотидов, транслокации, дупликации и инверсии различных частей гена. В том случае, когда под действием мутации изменяется лишь один нуклеотид, говорят о точковых мутациях. Поскольку в состав ДНК входят азотистые основания только двух типов — пурины и пиримидины, все точковые мутации с заменой оснований разделяют на два класса: транзиции (замена пурина на пурин или пиримидина на пиримидин) и трансверсии (замена пурина на пиримидин или наоборот). Возможны четыре генетических последствия точковых мутаций: 1) сохранение смысла кодона из-за вырожденности генетического кода (синонимическая замена нуклеотида), 2) изменение смысла кодона, приводящее к замене аминокислоты в соответствующем месте полипептидной цепи (миссенс-мутация), 3) образование бессмысленного кодона с преждевременной терминацией (нонсенс-мутация). В генетическом коде имеются три бессмысленных кодона: амбер — UAG, охр — UAA и опал — UGA (в соответствии с этим получают название и мутации, приводящие к образованию бессмысленных триплетов — например амбер-мутация), 4)обратная замена (стоп-кодона на смысловой кодон).
По влиянию на экспрессию генов мутации разделяют на две категории: мутации типа замен пар оснований и типа сдвига рамки считывания (frameshift). Последние представляют собой делеции или вставки нуклеотидов, число которых не кратно трем, что связано с триплетностью генетического кода.
Первичную мутацию иногда называют прямой мутацией, а мутацию, восстанавливающую исходную структуру гена, — обратной мутацией, или реверсией. Возврат к исходному фенотипу у мутантного организма вследствие восстановления функции мутантного гена нередко происходит не за счет истинной реверсии, а вследствие мутации в другой части того же самого гена или даже другого неаллельного гена. В этом случае возвратную мутацию называют супрессорной. Генетические механизмы, благодаря которым происходит супрессия мутантного фенотипа, весьма разнообразны.









Инверсия в пределах гена
Если инверсия участка ДНК происходит внутри одного гена, то такую мутацию относят к генным. Инверсии более крупных участков относятся к хромосомным мутациям.
Инверсия происходит вследствие поворота участка ДНК на 180°. Часто это происходит при образовании петли в молекуле ДНК. При репликации в петле репликация идет в обратном направлении. Далее этот кусок сшивается с остальной нитью ДНК, но оказывается перевернутым наоборот.
Если инверсия случается в смысловом гене, то при синтезе пептида часть его аминокислот будет иметь обратную последовательность, что скажется на свойствах белка.